

2 курс / Нормальная физиология / Общий_курс_физиологии_человека_и_животных_Том_1_Ноздрачев_А_Д_,
.pdfпередачи сенсорных сообщений, причем, чем большее число каналов расположено в пределах данной сенсорной системы, тем большее число таких переключений характерно для каждого из них.
Пути развития принципов многоканальности и многоуровневое™ в сенсорных системах иллюстрирует схема (рис. 4.4), из которой следует, что строение системы переключений обеспечивает и точную связь однозначных элементов на всех уровнях, и возможно более широкое взаимодействие между всеми элементами каждого уровня. Эти оба механизма необходимы для неискаженной передачи сведений об отдельных признаках и деталях наряду с объединением их при формировании целостного образа.
Рис. 4.4. Общая конструкция нейронных переключений в сенсорных системах. А — простая конструкция; Б — более сложная конструкция:1 — рецепторы, 2 — периферические чувствительные нейроны, 3 — низшая переключательная инстанция, 4 — средняя переключательная инстанция, 5 — высшая переключательная инстанция в коре
Однако установлено, что в пределах сенсорных систем существуют афферентные каналы срочной передачи информации (без переключений) в высшие мозговые центры. Считают, что по этим каналам осуществляется преднастройка высших мозговых центров к восприятию последующей информации. Появление таких каналов у высокоорганизованных животных и человека отражает общую тенденцию в совершенствовании конструкции мозга и повышении надежности его сенсорных систем.
В действительности надежность обеспечивается более сложным путем благодаря
частичному взаимному перекрытию нейронов. Концевые разветвления одного и того же нейрона подходят к не скольким нейронам следующего уровня (дивергенция, мультипликация). Но тот же нейрон может устанавливать контакты, получая импульсацию сразу от нескольких нейронов предыдущего уровня переключения (конвергенция) (рис. 4.5).
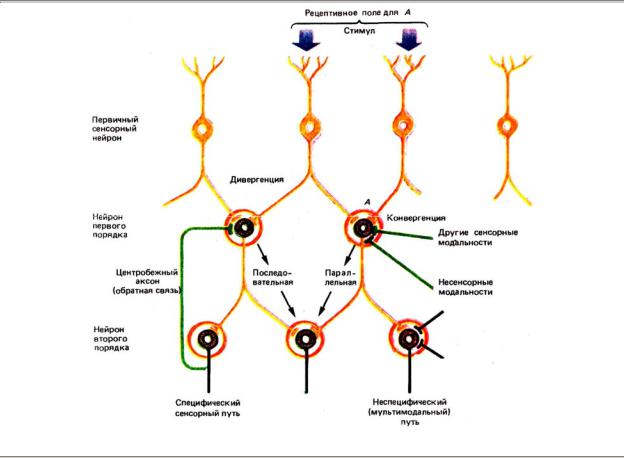
Рис. 4.5. Некоторые общие принципы организации сенсорных сетей
Таким образом, наличие ряда уровней в сенсорных системах, каждый из которых работает по принципу дивергенции и. конвергенции, выявляет возможность многомерной пространственной упорядоченности расположения нейронов, избирательно реагирующих на набор простых признаков сенсорных стимулов и поэтапную обработку информации.
Одной из общих черт организации сенсорных систем является принцип двойственной проекции их на кору больших полушарий. Этот принцип тесно связан с многоканальностью организации проводящих систем и заключается в существовании двух различных типов корковых проекций. В самой общей форме их можно разделить на первичные и вторичные проекции. Как правило, корковые проекции окружены ассоциативными корковыми зонами той же сенсорной системы.
Вкаждой сенсорной системе наряду с восходящими (афферентными) путями находятся
инисходящие (эфферентные, центрифугальные) пути. Восходящие и нисходящие волокна в сенсорных системах, переключаясь в одних и тех же мозговых центрах, теснейшим образом связаны друг с другом, что дает им возможность функционировать в надежном взаимодействии.
Наличие у различных стволовых и таламических образований сенсорных систем входов нисходящих путей указывает на то, что эти образования не следует рассматривать как просто релейные станции. В своей совокупности они представляют собой не пассивный канал линии связи, а являются аппаратами, где может осуществляться управление процессом передачи афферентной импульсации с нижележащих уровней на вышележащие.
Таким образом, существование нисходящих путей и их функциональная значимость позволяют признать наличие общего принципа обратных связей для всех сенсорных систем.
Нисходящие связи, по-видимому, выполняют роль «настроечной» функции, и данная функция должна быть тесно связана с аппаратами межсенсорной интеграции.
Рассмотрение процессов регуляции в сенсорных системах с точки зрения интегративной деятельности целого мозга позволяет представить нисходящие пути не только как систему внутрисенсорных связей, но и как систему общего конечного пути к рецепторным образованиям.
Учитывая многоканальность и многоуровневость организации любой сенсорной системы и существование специфического и ассоциативного каналов проведения информации, в пределах сенсорной системы можно выделить следующие уровни: рецепторный, стволовой, таламический и кортикальный. Оба канала связаны двусторонними взаимоотношениями внутри одного уровня и межуровневыми. Ассоциативный канал на всех его уровнях получает афферентацию от специфического канала и с помощью обратных связей может регулировать деятельность последнего.
Управляющая роль ассоциативных структур и функциональная значимость обратных связей при обработке афферентного потока свидетельствуют о том, что неотъемлемой частью процессов распознавания являются механизмы фильтрации. Последние возникают за счет целенаправленного взаимодействия прямых и обратных функциональных связей. При этом они обобщенно отражают комплекс функциональных операций, преобразующих информацию к некоторому виду, который обеспечивает дальнейшую и эффективную обработку.
Такое множество операций организуется за счет кортикального управления афферентным потоком в специфическом канале. Направленность подобного рода управляющих воздействий обеспечивается процессом избирательного внимания к некоторому набору биологически значимых признаков, характеризующих ситуацию.
Таким образом, вектор эволюции периферических и центральных звеньев сенсорных систем направлен на развитие способности животных получать все большую информацию об окружающей среде, все более подробно анализировать признаки сложных раздражителей, выявляя из них такие, которые выделяют адаптивное поведение и ориентацию в пространстве.
4.1.5. Сенсорное кодирование
Механические, химические, световые и другие раздражители, имея разнообразную природу и неся многостороннюю информацию об окружающей среде, преобразуются рецепторами в универсальные для мозга сигналы — нервные импульсы. В технике отображение одного сигнала другим принято называть кодированием. Следуя этой терминологии, можно говорить, что рецепторы кодируют информацию о среде, т. е. преобразуют сигналы, не воспринимаемые мозгом, в другие сигналы, «понятные» ему.
Какие же свойства раздражителя любой модальности способны преобразовывать и передавать рецепторные приборы в высшие центры? Прежде всего, это относится к качеству раздражителя, а затем к его количественным характеристикам: изменению интенсивности, временным показателям и пространственным признакам. Во всех этих преобразованиях участвуют не только рецепторы, но и следующие за ними цепи и центральные отделы нервной системы.
Кодирование качества. Очевидно, что различение действующих на организм внешних сил по их физической и химической природе происходит уже при первой встрече с ними соответствующих рецепторов. Это различение достигается избирательной чувствительностью рецепторов к определенной адекватной для них модальности раздражителя и исключительно низкими порогами возбуждения. Например, глаз раздражается светом, но не реагирует на звук, к которому чувствительно ухо, безразличное к свету. Однако встает вопрос, каким же образом мозг «узнает» модальность действующего стимула, т. е. распознает свет и звук, прикосновение и химическое воздействие? Если потенциалы действия всех нервных волокон в принципе одинаковы, то почему раздражители разных модальностей ощущаются не одинаково?
На первый взгляд ответ кажется очевидным — к каждому виду раздражителей приспособлены соответствующие рецепторы, связанные с вполне определенными отделами ЦНС. Однако в настоящее время установлено, что даже на уровне рецепторов в пределах одной и той же модальности существует своя специализация. Так, области поглощения трех цветовых пигментов в наружных сегментах колбочек у приматов хотя и перекрываются, но каждый рецептор имеет свои характерные максимумы. Или, например, вкусовые рецепторы способны в разной мере отвечать на различные по вкусовым качествам стимулы.
Таким образом, анализ отдельных признаков сенсорных стимулов в нервной системе не может быть основан на показании одного рецепторного элемента, а должен осуществляться их совокупной деятельностью.
Сенсорный проводящий путь состоит из ряда модально-специфических нейронов, соединенных синапсами. Такой принцип организации получил название меченой линии или топической организации. Этот принцип заключается в пространственно упорядоченном расположении нейронов на различных уровнях сенсорных систем соответственно характеристикам их рецептивных полей.
Понятие рецептивное поле имеет важное значение в сенсорной физиологии. С морфологической точки зрения, рецептивное поле — это тот участок рецепторной поверхности (например, кожи, сетчатки глаза и т. д.), с которым данная нервная структура (волокно, нейрон 1-го, 2-го, ..., n-го порядка) связана анатомически, т. е. жестко.
При функциональном подходе рецептивное поле — понятие динамическое, означающее, что один и тот же нейрон в различные отрезки времени в зависимости, например, от характеристики падающего раздражения, может оказаться связанным с различным числом рецепторов. Максимальная величина рецептивного поля нейрона в этом случае определяется его анатомическими связями, а минимальная — может ограничиваться всего одним рецептором.
Свойства рецептивных полей отражают возрастающую степень переработки информации и извлечения признаков, которое происходит в нейронах на все более высоких уровнях сенсорных путей.
Наличие дивергирующих и конвергирующих структурных связей могло бы вызывать обширнейшую иррадиацию возбуждения, тогда как слабый сенсорный стимул должен был бы запустить цепную реакцию нейронной сети всего мозга. Однако этого не происходит благодаря явлению латерального торможения, которое ограничивает иррадиацию возбуждения и тем самым обусловливает наличие рецептивных полей у центральных нейронов и, следовательно, топическую организацию сенсорных систем. Механизм латерального торможения основан на реципрокном тормозном взаимодействии между рецепторными элементами и между нейронами более высокого порядка.
За счет латерального торможения обеспечиваются избирательный характер реагирования нейронов при воздействии на определенные участки рецепторной поверхности и ее пространственно упорядоченная проекция в целом.
Принципу меченой линии до недавнего времени противопоставляли теорию «структуры ответа», согласно которой рецепторы кодируют качественные особенности раздражителей структурой импульсного ответа. Эта теория предполагала отсутствие жестких связей между рецепторами и соответствующими центральными нейронами. Основанием для нее послужило наблюдение, что кодирование информации осуществляется не одиночными импульсами, а группой равномерно следующих ПД. При этом появляется возможность использовать в качестве сигнальных признаков такие дополнительные параметры выходных сигналов, как частота импульсации или продолжительность межимпульсных интервалов.
Для неравномерно следующих импульсов сигнальными признаками могут служить число импульсов в «пачке» или продолжительность пачек, а также интервалы между ними и периодичность их следования. Возможности такого кодирования безграничны, так как
вероятны самые разнообразные вариации с пачками импульсов. Пространственновременные распределения электрической активности нервных волокон принято называть
паттернами.
Разнообразные качества стимулов, согласно этой теории, отображаются характерными «узорами» паттернов. Нейроны способны расшифровывать эти сигналы и в зависимости от их структуры формировать ощущения, которые соответствуют раздражителям, кодируемым определенными паттернами.
Нейрон, реагируя по-разному на различные паттерны, может участвовать в выполнении нескольких функций, причем каждый оттенок качества ощущений возникает в результате деятельности комплекса нейронов, образующих динамические ансамбли. Возникновение того или иного нейронного ансамбля зависит от характера паттернов, пришедших от рецепторов.
Различия в организации сенсорных систем по принципу меченой линии и в соответствии с теорией структуры ответа можно сравнить с путями развития телефонной связи. Прежде каждый провод телефонной сети связывал между собой лишь двух абонентов. Теперь по одному проводу могут вести переговоры десятки пар абонентов, так как сигналы на выходе телефона каждого абонента имеют свой код.
Экспериментальные исследования проблемы кодирования сенсорной информации привели к выводу, что невозможно объяснить всякое различение качества раздражителей лишь одной теоретической схемой. Для каждой основной модальности эволюция нашла свое решение в соответствии с физическими свойствами различаемых стимулов. Оказалось, что одни качества распознаются сенсорными системами, функционирующими по принципу топической организации, другие — кодируются паттернами. Так, распознавание многих качеств зрительных образов осуществляется мечеными линиями, а вкусовые раздражители кодируются паттернами.
Важной задачей, стоящей перед исследователями механизмов и способов кодирования информации рецепторными аппаратами, является количественная оценка информационных процессов.
Кодирование интенсивности. Количественный анализ раздражителей внешнего мира начинается со сравнительной оценки их интенсивности — более сильное раздражение вызывает более сильное ощущение. Так как частота афферентной импульсации зависит от амплитуды рецепторного потенциала, а она, в свою очередь, пропорциональна интенсивности раздражителя, то кодирование интенсивности стимула осуществляется посредством изменения частоты следования нервных импульсов от рецепторов в мозговые центры. Увеличение интенсивности раздражителя отображается в повышении частоты импульсной активности.
Как правило, между интенсивностью стимула и частотой потенциалов действия существует логарифмическая зависимость, т. е. ощущения увеличиваются пропорционально логарифму интенсивности раздражения. Это выражение получило название закона Вебера — Фехнера.
За счет логарифмического преобразования интенсивности рецепторы как бы сжимают информацию в нервных каналах связи, что обеспечивает высокую точность оценки интенсивности при малых ее изменениях. Предполагают, что логарифмическое преобразование информации в рецепторах облегчает ЦНС анализ и синтез сенсорной сигнализации.
Однако существует и другая точка зрения. Так, например, Стивене сформулировал закон степени, согласно которому интенсивность ощущения пропорциональна раздражению, возведенному в некую степень п, где для большинства ощущений значение п меньше единицы.
Однако при экспериментальной проверке оказалось, что и логарифмическая и степенная функциональные зависимости весьма сходны и могут быть справедливы для средней части рабочего диапазона того или иного органа.
Кроме того, способом кодирования интенсивности сенсорных стимулов является кодирование числом нервных элементов, участвующих в ответе. Этот способ кодирования имеет существенное значение, так как не все афференты обладают одним и тем же порогом возбуждения. Разница в порогах возбуждения обусловлена или различием генераторных потенциалов у рецепторов, или различием в легкости запуска механизма генерации импульсов, но так или иначе одни чувствительные волокна активируются при более слабых стимулах, чем другие.
Благодаря разнопороговым элементам зависимость количества рецепторов от интенсивности стимула будет иметь существенно большее значение для динамического диапазона, чем для одного нейрона. Есть основания полагать, что ЦНС «считывает» интенсивность по числу реагирующих элементов, умноженному на среднюю частоту импульсации.
Таким образом, изменения интенсивности раздражителя отображаются колебаниями частоты импульсации в отдельных афферентных волокнах и количеством каналов связи, по которым сигнализация поступает в мозг.
Пространственное кодирование. В некоторых сенсорных системах естественная стимуляция рецепторов характеризуется тем или иным распределением локальных стимулов. Способность определять место или конфигурацию стимулов называется
пространственным различением.
Наиболее простым способом воспроизведения пространственного расположения стимулов на рецептирующей поверхности является поточечное нанесение их на поверхность в ЦНС. Топическая организация сенсорных систем свидетельствует о многоканальной передаче сигнализации в высшие этажи мозга. Но каждый отдельный канал связи собирает информацию с более или менее широкой рецептивной области, а, кроме того, рецептивные поля смежных элементов часто перекрываются. Это перекрытие, во-первых, обеспечивает надежность функции, так как она всегда представлена большим числом элементов, а во-вторых, позволяет стимулам малой амплитуды вступать в контакт с более чувствительными рецепторами, а также вовлекать в возбуждение другие, менее чувствительные рецепторы.
В зрительной и слуховой системах удалось выделить афферентные каналы, пространственно разнесенные в центральных структурах и связанные с обработкой информации о локализации источника раздражения и его перемещения, а также о хроматических и частотных качествах сигнала.
Способность оценивать направление воздействия стимула присуща и самим рецепторам. Она выражается в том, что рецептор максимально возбуждается при каком-то определенном направлении действия стимула, а при другом не только не возбуждается, но даже тормозится. Например, импульсация в волосковых механорецепторах боковой линии рыб, обладающих в покое фоновой активностью, усиливается при движении жидкости в канале в одном направлении, вызывая деполяризацию, и угнетается при обратном токе, вызывая гиперполяризацию.
Чувствительность к направлению раздражения отмечена для тканевых механорецепторов (тельце Пачини), а также для электрорецепторов в различной реакции на действие катода и анода. Причинами направленной чувствительности рецепторов являются, по-видимому, особенности их микроструктуры, а также наличие полярной организации и строгой ориентации молекул воспринимающего субстрата.
Временное кодирование. Обсуждение оценки времени невозможно отделять от других аспектов кодирования. В самом общем виде частота нервных разрядов — это универсальная переменная величина, которая измеряется во времени. Кодирование информации, как правило, осуществляется группой равномерно следующих импульсов. При этом появляется возможность использовать в качестве сигнальных признаков такие временные параметры выходных сигналов, как частота импульсации или продолжительность межимпульсных интервалов. При неравномерно следующих
импульсах сигнальными признаками могут служить число импульсов в пачке или продолжительность пачек, а также интервалы между ними.
Для временного различения двух раздражителей также необходимо, чтобы вызванные ими нервные процессы не сливались во времени и чтобы сигнал, вызванный последующим стимулом, не попадал в рефрактерный период от предыдущего раздражения.
Помимо временных интервалов и длительности существуют и другие аспекты сенсорной стимуляции, связанные со временем. Они относятся к адаптационным процессам в рецепторах. Как было описано выше, адаптация определяет профиль ответа нервных элементов в зависимости от временной разномерности.
Таким образом, уже на уровне рецепторов осуществляется первичное кодирование качества стимулов и их количественных характеристик: переход из присущей им формы физической и химической энергии в форму нервных импульсов. Преобразованные сообщения поступают на следующий уровень сенсорной системы, где подвергаются дальнейшим преобразованиям, приводящим к изменению кода. Однако ни на одном из уровней сенсорной системы не происходит воссгановление стимула в его первоначальной форме, т. е. декодирование. Этим физиологическое кодирование отличается от большинства технических систем связи, где сообщение, как правило, восстанавливается в первоначальном, декодированном виде.
4.2. Соматическая сенсорная система
Кожа или любой другой наружный покров, отделяющий тело от внешней среды, существует у каждого организма. Через этот наружный покров и его производные животные получают информацию о физических изменениях среды, воздействиях со стороны других живых организмов и различных предметов. Покровы представляют собой сложные по структуре образования и выполняют многообразные функции: защитную, выделительную, обменную, дыхательную и др. Одной из них является сенсорная рецепция.
У беспозвоночных сенсорная функция покровов сводится в основном к дистантной тактильной рецепции механических раздражителей. Соматическая сенсорная система позвоночных животных в своем составе содержит три категории рецепторов, реагирующих на разные модальности раздражителей. Наиболее примитивной является болевая, или ноцицептивная, рецепция стимулов, которая сигнализирует о возможности повреждения организма. Вторая модальность — это способность чувствовать окружающую температуру. Восприятие этих раздражителей осуществляется терморецепторами. И, наконец, третьим видом рецепции покровов является
механорецепция, или тактильная чувствительность, реагирующая на разные виды механических стимулов (прикосновение, давление, вибрация и др.).
4.2.1. Соматическая сенсорная система беспозвоночных
Формирование всех типов животных происходило в водной среде. Эта среда обладает рядом специфических свойств, которые наложили отпечаток на развитие всех фундаментальных систем организмов, в том числе и на сенсорные. При движении объектов в водной среде возникают сложные физические явления. Ввиду малой сжимаемости воды вблизи движущегося объекта происходит смещение частиц среды — так называемый эффект ближнего поля.
Мелкие подвижные организмы, обладающие ритмически работающими органами (мембраны, жгутики, реснички, плавники и др.), в силу большой амплитуды колебаний и высокой плотности воды являются эффективными источниками ближнего поля. Отсюда становится очевидным, что уже первые живые организмы сталкивались с эффектами
ближнего поля и могли выработать рецепторную систему для их восприятия, чтобы успешно ориентироваться в среде, населенной другими организмами. Способность обнаруживать подвижные объекты на расстоянии создавала явные преимущества в пищевом в оборонительном поведении, и неудивительно, что такая рецепторная система возникла уже на самых ранних этапах эволюции. Она свойственна уже бактериям и простейшим.
Хорошо известны реакция реверсии (изменение на противопо-ложное) направления движения бактерий при приближении к препятствию или направленное перемещение по (или против) градиенту силы тяжести. У инфузорий слабое тактильное раздражение вызывает местную реакцию реверсии биения ресничек, а при большой интенсивности раздражения возникает направленная реакция избегания стимула. Эта реакция может быть вызвана при раздражении любой части тела простейшего. Реакция на тактильную стимуляцию связана у них с первичной механочувствительностью всей поверхностной мембраны.
При механической стимуляции поверхности мембраны простейших возникает градуальное деили гиперполяризационное изменение мембранного потенциала, распространяющееся во все стороны с декрементом, которое можно рассматривать как РП. При достижении определенной амплитуды рецепторный потенциал переходит в спайковый, распространяющийся без декремента и вызывающий реверсию биения ресничек или сокращения тела. Генерация спайков происходит в любом участке тела клетки, где амплитуда РП достигла надпорогового уровня. Предполагают, что первичные механорецепторные процессы приводят к высвобождению ионов кальция, в покое связанных мембранным матриксом, которые активируют АТФазную систему актомиозиновых комплексов сократительных органелл.
У многоклеточных беспозвоночных животных, начиная с кишечнополостных, уже существует специализированная сенсорная система, способная к восприятию смещений ближнего поля. В ходе эволюции она совершенствуется в связи с необходимостью обеспечения пищевого и оборонительного поведения или ориентации по отношению к водным и воздушным потокам.
Гидроидные полипы реагируют на проплывающих мимо рачков изгибанием тела и щупалец в направлении добычи и последующим ее захватом. В этом случае стимулом служат низкочастотные смещения частиц воды в ближнем поле, к которым полипы очень чувствительны. Пороговая величина смещения составляет всего 2—3 мкм. Рецепторами являются гиподермальные чувствительные клетки, несущие на дистальном конце длинную ресничку (рис. 4.6). Эти клетки расположены диффузно на поверхности щупалец. Реснички торчат наружу и непосредственно подвергаются действию внешнего стимула. Эти рецепторы еще не имеют нейрита и, по-видимому, передают возбуждение прямо на соседние эпителиально-мышечные клетки. В ответ на раздражение гиподермальные рецепторы генерируют лишь градуальный РП, а спайковый ответ возникает только в нервной сети в результате суммации сигналов рецепторов.
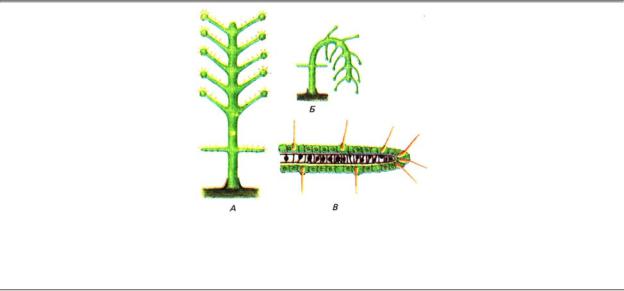
Рис. 4.6. Дистантные механорецепторы гидроидных полипов. А — гидроидный полип с пятью ярусами булавовидных щупалец и одним ярусом нитевидных щупалец; Б — реакция изгибания полипа в сторону механического раздражения одного из щупалец; В — продольный разрез через верхушку нитевидного щупальца (видны торчащие наружу реснички механорецепторов)
Впервые у плоских червей происходят окончательная дифференциация гиподермальных рецепторов на хемо- и механорецепторы. В свою очередь, механорецепторы подразделяются на контактные тактильные и дистантные рецепторы, предназначенные для восприятия водных смещений. Эти последние несут на дистальном конце длинные немоторные реснички, далеко выступающие за уровень ресничек покровного эпителия. Сома рецепторов лежит глубоко под эпителием, а нейрит уходит в ЦНС. Как и у кишечнополостных, между соседними рецепторами существует прямое взаимодействие на уровне нервного сплетения. У кольчатых червей — полихет —
вгиподерме обнаружено несколько типов сенсорных образований, которые по своей морфологии могут рассматриваться как механорецепторы.
Учленистоногих основным биохимическим компонентом кутикулы служит хитин. Так как в кутикуле кольчатых червей хитина нет, считают, что его появление у членистоногих представляет значительный эволюционный шаг и служит важной биохимической меткой при изучении эволюции. У большинства членистоногих хитиновые покровы жестки, что придает необходимую твердость экзоскелету, но требует и соответствующих приспособлений сенсорных рецепторов для обнаружения механических и других стимулов.
Простым тактильным рецептором может служить трихоидная сенсилла, обслуживаемая одним биполярным нейроном (рис. 4.7). Воспринимающая часть сенсиллы представлена волоском — пустотелым выростом кутикулы, подвижно сочлененным с покровами с помощью сочленовной мембраны. К его основанию подходит дендрит биполярного нейрона, на кончике которого расположен специализированный цилиарный аппарат в виде одной или нескольких подвижных ресничек. Как и в других типах рецепторов, через этот аппарат осуществляется преобразование энергии внешнего стимула в электрическую энергию первичного импульса. При отклонении волоска в предпочтительном направлении
вчувствительном нейроне возникает деполяризационный РП, а при отклонении в противоположную сторону — гиперполяризационный. Когда РП достигает критического уровня, в основании аксона возникают распространяющиеся бездекрементно потенциалы действия.
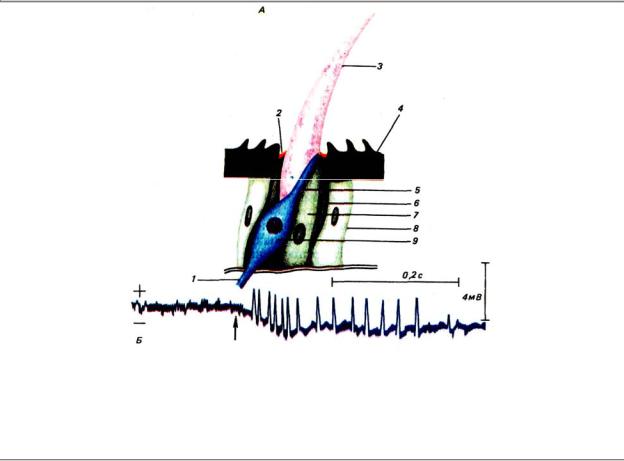
Рис. 4.7. Строение механорецепторной трихоидной сенсиллы (А) и ее электрический ответ на изгиб волоска (Б): 1 — аксон рецепторного нейрона, 2 — сочленовная мембрана, 3 — волосок, 4 — кутикула, 5 — дендрит, 6 — клетка, образующая сочленовную мембрану, 7 — трихогенная клетка, 8 — клетка эпидермиса, 9 — нейрон; на осциллограмме регистрируются медленный рецепторный потенциал (отклонение основной линии вниз) и быстрые колебания потенциала действия; стрелкой обозначено начало раздражения
Кроме нейрона каждая сенсилла включает еще три эпителиальные клетки, окружающие рецептор. Появление дополнительных клеток, взявших на себя часть функций, которые у низших беспозвоночных выполнялись самим рецепторным нейроном, несомненно, прогрессивное явление. Оно обеспечило большую автономность рецепторных единиц и создало возможность для более глубокой и пластичной специализации каждого из компонентов сенсиллы.
Разбросанные по всему телу насекомых однонейронные трихоидные сенсиллы выполняют функцию осязательных рецепторов. Любая сила, смещающая волосок, — прикосновение, движение воздуха или воды, изменение давления — стимулирует сенсорную клетку. Сенсорные клетки в кутикуле поверхности тела посылают волокна к соответствующему сегментарному ганглию нервной цепочки.
Таким образом, параллельно с развитием рецепторного отдела механорецепторных сенсилл возникают специализированные центрольные нервные элементы для независимой обработки сигналов от каждого из видов рецепторов. Развиваются центры, формирующие разнообразные и значительно более тонкие реакции организма в соответствии с требованиями окружающей среды, чем это наблюдалось у низших беспозвоночных. Центральные проекции рецепторов становятся более детерминированными, т. е. менее диффузными.
Следовательно, эпидермальные рецепторы перестают работать как диффузная чувствительная нервная система, а функционируют теперь как дискретные рецепторные органы. Они связаны с нервными структурами, ответственными за контроль определенных поведенческих реакций целостного организма.
4.2.2.Соматическая сенсорная система позвоночных
Вотличие от беспозвоночных животных все позвоночные лишены кутикулы и их тела