

2 курс / Нормальная физиология / Общий_курс_физиологии_человека_и_животных_Том_1_Ноздрачев_А_Д_,
.pdfборозды. К неоцеребеллуму по трактам, переключающимся в ядрах варолиева моста, поступает афферентная импульсация от обширных областей коры больших полушарий (лобных, теменных, височных и затылочных долей).
Полушария и червь мозжечка состоят из лежащего на периферии серого вещества — коры — и расположенного глубже белого вещества, в котором заложены скопления нервных клеток, образующие ядра мозжечка. Кора мозжечка представлена тремя слоями, каждый из которых имеет определенный набор клеточных элементов. Самый поверхностный слой — молекулярный — состоит из параллельных волокон и разветвлений дендритов и аксонов нейронов нижележащих слоев. В нижней части молекулярного слоя расположены тела корзиночных клеток, аксоны которых оплетают тела и начальные сегменты аксонов клеток Пуркинье (рис. 3.11, Б). Здесь же в молекулярном слое имеется некоторое количество звездчатых клеток.
Вентральнее молекулярного слоя находится ганглиозный слой, в котором сосредоточены тела клеток Пуркинье (рис. 3.11, Б). Эти крупные клетки ориентированы вертикально по отношению к поверхности коры мозжечка. Их дендриты поднимаются вверх и широко ветвятся в молекулярном слое. Дендриты клеток Пуркинье содержат множество шипиков, на которых образуют синапсы параллельные волокна молекулярного слоя. Аксоны клеток Пуркинье спускаются к ядрам мозжечка. Часть на них заканчивается на вестибулярных ядрах. Практически аксоны клеток Пуркинье представляют собой
единственный выход из коры мозжечка.
Под ганглиозным слоем лежит гранулярный слой, который содержит большое число тел клеток-зерен, или гранулярных клеток. По некоторым подсчетам их число может достигать 10 млрд. Аксоны клеток-зерен поднимаются вертикально вверх в молекулярный слой и там Т-образно ветвятся. Ветви идут параллельно поверхности коры и образуют синапсы на дендритах других клеток. Здесь же в гранулярном слое лежат клетки Гольджи, аксоны которых подходят к клеткам-зернам.
Афферентный вход к нейронному аппарату коры осуществляется по трем системам волокон. Это, во-первых, лазающие, или лиановидные, волокна, идущие из нижних олив продолговатого мозга. Нижняя олива получает афференты от нескольких восходящих трактов спинного мозга и из центров головного мозга. Лазающие волокна широко ветвятся и подобно лианам оплетают дендриты клеток Пуркинье, формируя на них синапсы. Вторая система афферентных волокон — это мшистые, или моховидные, волокна, идущие от ядер моста и оканчивающиеся на клетках-зернах. Мшистые волокна многократно ветвятся и образуют синапсы на множестве клеток коры мозжечка. И, наконец, третья система афферентных волокон — это также широко ветвящиеся адренэргические волокна, поступающие в кору мозжечка из голубого пятна в среднем мозгу. Голубое пятно представляет собой скопление из нескольких сотен нейронов, аксоны которых способны диффузно выбрасывать норадреналин в межклеточное пространство. Вероятно, эти нейроны выполняют нейромодуляторную функцию и могут изменять возбудимость нейронов, локализованных в коре мозжечка.
Нейрофизиологические исследования Дж. Экклса показали, что корзинчатые и звездчатые клетки, которые заканчиваются синапсами на клетках Пуркинье, вызывают в них тормозные постсинаптические потенциалы (ТПСП) и подавление импульсной активности. Клетки Гольджи тормозят клетки-зерна по принципу обратной связи (рис. 3.11, Б).
Таким образом, большинство связей, опосредованных интернейронами коры мозжечка, являются тормозными. Исключение составляют только клетки-зерна, которые возбуждаются от мшистых волокон и сами через Т-образно ветвящиеся аксоны активируют все остальные интернейроны коры мозжечка. Однако конечный эффект этой активации опять-таки сводится к торможению.
Клетки Пуркинье, которые представляют собой выход функциональной системы, могут возбуждаться прямо через лиановидные волокна и опосредованно через моховидные
волокна и клетки-зерна. Возникающие под действием этого возбуждения разряды клеток Пуркинье, согласно электрофизиологическим данным, вызывают в конечном итоге торможение нейронов ядер мозжечка. Эти факты свидетельствуют о том, что деятельность всей нейрональной системы коры мозжечка сводится к торможению ядер, над которыми кора надстроена. Очевидно, механизм этого торможения можно представить следующим образом,
Впокое клетки Пуркинье обладают фоновой электрической активностью, которая вызывает тоническое торможение нейронов в ядрах мозжечка. Возбуждение клеток Пуркинье через систему лиановидных или мшистых волокон приводит к увеличению частоты импульсных разрядов этих нейронов и, как следствие, к усилению торможения ядер мозжечка. Напротив, торможение клеток Пуркинье, вызванное звездчатыми или корзинчатыми клетками, сопровождается растормаживанием нейронов в ядрах мозжечка. Сами же ядра мозжечка, обладающие постоянной тонической активностью, через нисходящие пути регулируют уровень возбудимости центров спинного мозга и мышечный тонус.
Согласно гипотезе, высказанной Дж. Экклсом, большое количество тормозных нейронов в коре мозжечка предотвращает длительную циркуляцию возбуждения по нейронным цепям. Любой возбуждающий импульс, приходя в кору мозжечка, превращается в торможение за время порядка 100 мс. Так происходит как бы автоматическое стирание предшествующей информации, которое позволяет коре мозжечка участвовать в регуляции быстрых движений.
Вбелом веществе мозжечка сконцентрированы три пары ядер. В белом веществе червя близко к срединной плоскости находится ядро шатра, или фастигиальное ядро. Нейроны этого ядра посылают свои отростки к вестибулярному ядру Дейтерса и к ретикулярной формации продолговатого мозга и варолиева моста, где берет свое начало цетикулоспинальный тракт спинного мозга. Латеральнее фастигиального ядра находится
вставочное, или промежуточное, ядро, которое у человека разделяется на шаровидное и пробковидное ядра. От вставочного ядра аксоны идут в средний мозг к красному ядру. Менее развитый афферентный путь от вставочного ядра идет в промежуточный мозг к вентролатеральному ядру зрительного бугра — таламуса (см. разд. 3.7.2) — и оттуда к двигательной коре. Латеральнее всех ядер лежит наиболее крупное зубчатое ядро мозжечка, от которого мощные пучки волокон направляются к вентролатеральному ядру таламуса, и далее аксоны нейронов второго порядка проецируются в моторные зоны коры.
Кнейронам мозжечковых ядер подходят аксоны клеток Пуркинье. Установлено, что клетки Пуркинье червя устанавливают прямые связи с ядром Дейтерса. Это позволяет иногда относить ядро Дейтерса к внутримозжечковым ядрам по функциональному принципу.
Существует определенная топография связей коры мозжечка с его ядрами. Согласно классификации Бродала, кору мозжечка млекопитающих можно разделить на три продольные зоны: медиальную червячную зону, от которой аксоны клеток Пуркинье проецируются на ядро шатра, промежуточную зону коры, связанную со вставочным ядром, и латеральную зону коры полушарий, дающую проекции к зубчатому ядру. Эта классификация, в основу которой положены эфферентные связи мозжечка, свидетельствует о том, что латеральные отделы мозжечка через зубчатое ядро связаны с более высокими уровнями головного мозга.
Вцелом мозжечок имеет обширные эфферентные связи со всеми двигательными системами стволовой части мозга: кортикоспинальной, руброспинальной, ретикулоспинальной и вестибулоспинальной. Не менее разнообразными являются и афферентные входы мозжечка.
Афферентная информация в мозжечок от спинного мозга приходит по спинномозжечковым трактам (дорсальному и вентральному), ростральному спинномозжечковому и кунеоцеребеллярным трактам, по спинооливомозжечковым путям. Кора
больших полушарий также посылает афферентные пути в мозжечок, среди которых наиболее важными являются кортикоретикуломозжечковый и це-ребромостомозжечковый тракты.
3.6.2. Функции мозжечка
Мозжечок как надсегментарный орган, входящий в систему регуляции движений, выполняет следующие важные функции: 1) регуляция позы и мышечного тонуса; 2) сенсомоторная координация позных и целенаправленных движений; 3) координация быстрых целенаправленных движений, осуществляемых по команде из коры больших полушарий. Эти функции мозжечка наиболее удобно разбирать в соответствии с топической классификацией его отделов, основанной на характере эфферентных связей.
Медиальная червячная зона мозжечка в наибольшей степени связана с реализацией первой функции, т. е. с управлением тонуса, позы и равновесия тела. В эту зону коры мозжечка, а также в флоккулонодулярную долю архицеребеллума поступает афферентная информация, сигнализирующая о позе и состоянии локомоторного аппарата. После обработки этой информации из коры мозжечка через ядро шатра коррегирующие команды направляются к вестибулярному ядру Дейтерса, к ретикулярной формации ствола и оттуда к спинальным центрам по ретикулоспинальным и вестибулоспинальным трактам.
Таким образом, вся система работает по принципу обратной связи и обеспечивает срочную регуляцию антигравитационного тонуса.
Удаление червячной зоны мозжечка вызывает растормаживание ядра Дейтерса и как результат — усиление тонуса разгибателей у животного в состоянии децеребрационной ригидности. Электрическое раздражение червячной зоны, напротив, приводит к ослаблению тонуса мышц разгибателей за счет активации тормозных влияний мозжечка на ядро Дейтерса.
Промежуточная зона коры мозжечка, связанная со вставочным ядром (или шаровидным и пробковидным), так же как и предыдущая зона, имеет афферентные входы от спинальных трактов. Однако, кроме того, через ядра моста промежуточная зона коры мозжечка получает информацию от двигательной области коры больших полушарий. Эта информация поступает через коллатерали кортикоспинального тракта и сигнализирует о готовящемся целенаправленном движении. Сопоставление приходящей по этим двум путям информации позволяет промежуточной зоне мозжечка участвовать в координации целенаправленных движений с рефлексами поддержания позы, в выборе наиболее оптимальной для выполнения движения позы. Нисходящие команды от промежуточной зоны коры мозжечка через вставочное ядро идут к красному ядру и далее по руброспинальному тракту к моторным центрам спинного мозга.
Как уже отмечалось выше, латеральная, филогенетически наиболее молодая, кора полушарий мозжечка дает эфферентные проекции к зубчатому ядру. Эта латеральная зона через церебромостомозжечковый тракт получает афферентный вход от различных ассоциативных зон коры больших полушарий. По этим афферентным путям в кору полушарий мозжечка поступает информация о замысле движения. В полушариях и зубчатом ядре мозжечка эта информация преобразуется в программу движения, которая по мозжечковоталамокортикальному тракту поступает в двигательные зоны коры больших полушарий. В дальнейшем двигательный акт реализуется за счет нисходящих команд, идущих от двигательной коры в спинной мозг по кортикоспинальному тракту. Кроме того, от зубчатого ядра мозжечка через красное ядро могут распространяться прямые нисходящие воздействия к спинальным центрам.
Эта сложная система регуляции движений с наличием обратных связей между мозжечком и корой больших полушарий позволяет полушариям мозжечка участвовать в
организации быстрых целенаправленных движений, протекающих без учета информации,
идущей от восходящих сенсорных путей спинного мозга. Такие движения могут
встретиться в спортивной практике, при игре на музыкальных инструментах и при некоторых других видах деятельности.
Экспериментальные исследования функций мозжечка показали, что этот отдел головного мозга участвует не только в регуляции движений, но и включен в систему контроля висцеральных функций. Л. А. Орбели было установлено, что раздражение мозжечка вызывает целый ряд вегетативных рефлексов, таких, например, как расширение зрачков, повышение артериального давления и т. д. Удалению мозжечка сопутствуют нарушения сердечно-сосудистой деятельности, дыхания, моторики и секреторной функции желудочно-кишечного тракта. Электрофизиологическими методами в мозжечке обнаружены висцеральные проекции внутренних органов. Так, например, при раздражении интерорецепторов в коре мозжечка регистрируются вызванные потенциалы.
Все эти экспериментальные факты позволили Л. А. Орбели считать мозжечок млекопитающих не только регулятором моторной деятельности, но и адаптационно-
трофическим органом, который определяет возбудимость вегетативных и соматических центров. Однако до сих, пор остается неясным вопрос о том, какое место занимает мозжечок в иерархии структур, регулирующих висцеральные функции. По всей вероятности, модулирующие влияния мозжечка могут быть опосредованы связями с ретикулярной формацией СТВОЛЕ и гипоталамусом, который является главным подкорковым центром регуляции висцеральных функций.
Основные функции мозжечка определяют и характер патологических симптомов при нарушении его деятельности. Согласно классическим представлениям Л. Лючиани, сформулированным еще в конце прошлого столетия, при частичном и общем поражении мозжечка наблюдается три симптома: атония, астения и астазия.
Атония характеризуется ослаблением мышечного тонуса. У собак и кошек после удаления мозжечка наблюдается начальное повышение тонуса мышц-разгибателей, причем животные находятся в позе, сходной с состоянием децеребрационной ригидности. Однако через несколько суток после операции у животных гипертонус разгибателей сменяется гипотонией. Аналогичная операция у приматов сразу вызывает падение мышечного тонуса и, как следствие, неспособность поддерживать определенную позу. Обычно атония сопровождается симптомом астении, которая характеризуется слабостью и быстрой утомляемостью мышц. Очевидно, атония и астения связаны в первую очередь с выпадением функции мозжечка контролировать познотонические рефлексы.
Третий симптом, описанный Л. Лючиани, — астазия — проявляется в способности мышц давать колебательные и дрожательные движения. Мышечный тремор особенно выражен в начале и конце движения, что в значительной мере препятствует завершению целенаправленного движения. Этот симптом вызывается нарушением сенсомоторной координации позных и целенаправленных движений.
Более поздние исследования, проведенные на млекопитающих, показали, что удаление мозжечка пагубно сказывается на выполнении произвольных движений. При этом более всего страдают содружественные движения — симптомы асинергии. В результате асинергии происходит как бы распад программы движения, и целостное движение состоит не из одновременных содружественных актов, а из последовательности ряда простых движений. Так, например, касание рукой кончика носа мозжечковый больной осуществляет в три приема. Асинергия сочетается с дисметрией, или утратой соразмерности движения. Движения становятся преувеличенно размашистыми и теряют свою точность, в результате чего цель достигается только после многократных ошибок. У мозжечковых больных деформируется походка — атаксия. Атаксическая походка характеризуется широко расставленными ногами и избыточными движениями, из-за которых больного как пьяного «бросает» из стороны в сторону.
Нарушение координации быстрых целенаправленных движений при удалении или травмах мозжечка может вызвать адиадохокинез — неспособность выполнять быструю последовательность движений, например сгибание и разгибание пальцев.
Следует отметить, что у млекопитающих, перенесших травму мозжечка, со временем наступает довольно эффективная компенсация его функций. По всей вероятности, эта компенсация осуществляется за счет функции коры больших полушарий, которая обладает двусторонними связями с мозжечком. Этот факт свидетельствует о высокой пластичности мозговых систем регуляции движений.
3.7.1. Структура промежуточного мозга
Промежуточный мозг в процессе эмбриогенеза развивается из переднего мозгового пузыря и образует стенки третьего мозгового желудочка. Топографически и функционально промежуточный мозг подразделяется на эпиталамус, таламус и гипоталамус.
Эпиталамус, или надталамическая область, состоит из расположенного под мозолистым телом свода и из железы внутренней секреции эпифиза, которые формируют верхнюю стенку третьего желудочка. Таламус, или зрительный бугор, представляет собой состоящее из скопления серого вещества объемистое тело яйцевидной формы. Нижней и латеральной поверхностью таламус сращен с соседними частями мозга. Медиальная поверхность зрительного бугра образует боковую стенку полости третьего желудочка. Таламус является крупным подкорковым образованием, через которое в кору больших полушарий проходят разнообразные афферентные пути.
Дно третьего желудочка формирует группа структур, которые объединяют под названием гипоталамуса или подбугорья. Гипоталамус содержит большое количество ядер и является центром регуляции висцеральных функций организма (см. разд. 3.7.3).
3.7.2. Морфофункциональная организация таламуса
Нервные клетки таламуса группируются в большое количество ядер (до 40), которые топографически разделяют на переднюю, заднюю, срединную, медиальную и латеральную группы. По функции таламические ядра можно дифференцировать на специфические, неспецифические, ассоциативные и моторные.
В специфических, или проекционных, ядрах таламуса происходит синаптическое переключение сенсорной информации с аксонов восходящих афферентных путей на следующие, конечные нейроны, отростки которых идут в соответствующие сенсорные проекционные области коры больших полушарий. Повреждение специфических ядер приводит к необратимому выпадению определенных видов чувствительности. Эти экспериментальные факты свидетельствуют о том, что специфические ядра являются передаточной станцией на пути афферентных импульсов от периферических рецепторов к коре больших полушарий.
Среди основных проекционных ядер таламуса можно выделить вентробазальное ядро, которое является специфическим ядром соматосенсорной системы. Оно разделяется на две части — вентральное постериолатеральное ядро, к которому подходят восходящие волокна спиноталамического тракта и системы медиальной петли, несущие информацию от кожных рецепторов туловища, проприоцепторов мышц и суставного аппарата, и вентральное постериомедиальное ядро, к которому подходят соответствующие пути от ядер тройничного нерва, осуществляющего иннервацию лицевой части головы.
Микроэлектродные исследования нейронов вентробазального комплекса показали, что данное ядро, как и прочие специфические ядра, организовано по топическому принципу. Суть этого принципа состоит в том, что каждый нейрон активируется раздражением рецепторов определенного участка кожи, причем смежные участки туловища проецируются на смежные части вентробазального комплекса.
Специфичность данных нейронов проявляется также в том, что каждый из них возбуждается одним типом рецепторов. Соматотопический принцип организации
сохраняется и на более высоком уровне в соматосенсорной проекционной области коры больших полушарий (постцентральная извилина), с которой вентробазальный комплекс связан восходящими и нисходящими путями.
Специфическим ядром зрительной сенсорной системы является латеральное коленчатое тело (ЛКТ), имеющее прямые связи с затылочными (зрительными) проекционными областями коры больших полушарий.
Латеральное коленчатое тело имеет слоистую структуру и организовано также по топическому принципу. Аксоны, идущие в ЛКТ из зрительного тракта, распределяются в нем с поразительной четкостью: три слоя ЛКТ связаны с ипсилатеральным глазом, а три остальные — с контралатеральным. В каждом из слоев ЛКТ аксоны зрительного тракта вступают в синаптический контакт с четко ограниченными группами клеток. Нейроны ЛКТ обладают концентрическими рецептивными полями, центр и периферия которых антагонистичны и по-разному реагируют на изменение освещенности. Рецептивное поле нейрона ЛКТ представляет собой концентрически организованную совокупность рецепторов сетчатки, имеющую либо возбуждающий (при включении света) центр и периферическую тормозную часть, либо, напротив, тормозный центр и возбуждающую данный нейрон ЛКТ периферическую область.
Такая организация рецепторного поля позволяет нейронам ЛКТ хорошо реагировать на контраст при определении границы между темным и светлым и на суммарную яркость светового стимула.
У млекопитающих некоторые нейроны ЛКТ обладают цветоспецифичными рецепторными полями и могут возбуждаться или, наоборот, тормозиться в зависимости от длины волны светового стимула. Таким образом, нейроны ЛКТ, также как и нейроны сетчатки, принимают участие в анализе зрительной информации. Об этом же свидетельствует наличие тормозных процессов в нейронах ЛКТ, субстратом которых являются цепи отрицательной обратной связи, образованные возвратными коллатералями аксонов этих клеток и промежуточными нейронами, имеющими тормозные окончания. По всей вероятности, возникающие в нейронах ЛКТ тормозные постсинаптические потенциалы служат для подавления следовых эффектов и всех побочных воздействий, которые мешают передаче сигнала через данный канал.
Восходящие пути слуховой системы, идущие из нижних бугров четверохолмия и по волокнам латеральной петли, проецируются в специфическое таламическое ядро — медиальное коленчатое тело (МКТ), от которого начинается тракт, достигающий первичной слуховой коры в верхней части височных долей.
Медиальное коленчатое тело состоит из мелкоклеточной и крупноклеточной частей и обладает тонотопической специализиацией своих нейронов. Так, например, нервные клетки мелкоклеточной части МКТ имеют довольно узкую настройку на восприятие звуков различной высоты и принимают участие в анализе и передаче акустической информации.
Таким образом, по справедливому выражению одного из исследователей таламуса А. К. Уолкера, «таламус является посредником, в котором сходятся все раздражения от внешнего мира и, видоизменяясь здесь, направляются к подкорковым и корковым центрам...».
В специфические ядра таламуса проецируются афференты не только от экстерорецепторов и рецепторов двигательного аппарата. Электрофизиологические исследования показали, что в вентробазальном комплексе таламуса имеются области проекций блуждающего и чревного нервов, чувствительные волокна которых несут информацию от интероцепторов. В то же время таламус, как надсегментарный центр рефлекторной деятельности, имеет связи с гипоталамусом, где сосредоточены главные вегетативные центры. Эти связи характерны для передней группы ядер таламуса и создают материальную предпосылку для участия этой структуры в системе регуляции висцеральных функций организма.

Следующую функциональную группу ядер таламуса составляют гак называемые ассоциативные ядра. В отличие от специфических ядер они не могут быть отнесены к какой-либо одной сенсорной системе и получают афферентные импульсы от специфических проекционных ядер. Три ядра этой группы имеют связи с главными ассоциативными областями коры: ядро подушки связано с ассоциативной зоной теменной и височной коры, заднее латеральное ядро — с теменной, корой, медиальное дорсальное ядро — с лобной долей. Четвертое ядро — переднее — имеет связи с лимбической корой больших полушарий. По-видимому, ассоциативные ядра участвуют в высших интегративных процессах, однако их функция изучена еще недостаточно.
К моторным ядрам таламуса относится вентролатеральное ядро, которое имеет вход от мозжечка и базальных ганглиев и одновременно дает проекции в моторную зону коры больших полушарий. Это ядро включено в систему регуляции движений, и, как показал материал клиники, разрушение некоторых его участков ослабляет симптомокомплекс болезни Паркинсона.
Наконец, последнюю большую группу ядер таламуса образуют неспецифические ядра, которые функционально связаны с ретикулярной формацией ствола. К числу этих ядер относится срединная и интроламинарная группа ядер таламуса, которая получает афферентный вход от волокон, восходящих из ретикулярной формации, и, кроме того, Имеет двусторонние связи со специфическими ядрами таламуса. В отличие от специфических ядер с локальными проекциями в коре филогенетически более древние неспецифические ядра обнаруживают диффузные проекции во все области коры (рис. 3.12). Этой структурной особенностью обусловлены их название и функция, которая состоит в регуляции возбудимости и электрической активности корковых нейронов.
Рис. 3.12. Проекции специфических (а) и неспецифических (б) ядер таламуса в кору больших полушарий: 1 — сенсорные афферентные пути, 2 — ретикулоталамические пути, 3 — проекционная зона коры, 4 — ассоциативная зона коры
Доказательства влияния неспецифических ядер таламуса на кору впервые были получены американскими исследователями Э. Демпси и Р. Моррисоном в 1942 г. Они показали, что при электрическом раздражении неспецифических ядер с ритмом 6—12 имп/с почти на всей поверхности коры ипсилатерального полушария регистрируются постепенно увеличивающиеся по амплитуде негативые волны, очень сходные с альфаритмом электроэнцефалограммы. Эти негативные волны появляются с латентным периодом 25 мс и более на 2—3-й стимул, достигают максимальной амплитуды на 5—6-й стимул, а затем начинают постепенно уменьшаться вплоть до полного исчезновения. Если раздражение продолжается, то негативные волны появляются вновь в такой же последовательности. Эта электрофизиологическая реакция была названа рекруитирующим ответом или реакцией вовлечения.
В отличие от первичных ответов, которые регистрируют при раздражении специфических ядер, реакция вовлечения характеризуется большим латентным периодом,
нарастанием и снижением амплитуды и отсутствием локальности или диффузностью вне связи с какой-либо специфической областью коры.
Дальнейший анализ этого электрофизиологического феномена показал, что периодическое увеличение и уменьшение негативных волн реакции вовлечения обусловлено лучшей или худшей синхронизацией активности таламических и корковых нейронов, а также суммацией постсинаптических потенциалов в большем или меньшем количестве нейронов. Уровень синхронизации может определяться динамикой возбуждающих и тормозных процессов при таламокортикальных циклических взаимодействиях.
Наличие этих взаимодействий используют для объяснения природы ритмической активности коры больших полушарий и, в частности, происхождения альфа-ритма электроэнцефалограммы (см. разд. 3.11.1).
При сопоставлении функций специфических и неспецифических ядер таламуса возникает резонный вопрос о взаимодействии этих двух систем, которые могут влиять на одни и те же нейроны коры больших полушарий. Как показали электрофизиологические исследования, восходящие влияния неспецифических ядер таламуса проявляются не в вызове разряда коркового нейрона, а в изменении его возбудимости. Неспецифические влияния из таламуса, повышая возбудимость корковых нейронов, облегчают их деятельность, при этом ответы корковых нейронов на импульсы, приходящие из специфических проекционных ядер, усиливаются. Вместе с тем неспецифические влияния могут иметь и противоположный знак и обнаруживать угнетающее действие на разряды корковых нейронов.
Существует точка зрения, что неспецифические ядра включены в восходящую активирующую систему и являются посредниками между корой и ретикулярной формацией ствола, которая получает информацию от всех органов чувств. Таким образом, неспецифические ядра передают активирующие влияния ретикулярной формации и участвуют в поддержании оптимального тонуса коры. Однако эта точка зрения не является общепризнанной, и некоторые исследователи рассматривают ретикулярную формацию и неспецифические ядра таламуса как две раздельные системы, контролирующие возбудимость корковых нейронов.
Обсуждая таламокортикальные взаимодействия, было бы упрощением рассматривать их как односторонние. Кора, в свою очередь, может оказывать тормозные и облегчающие влияния на таламические ядра. Эти влияния могут распространяться на специфические ядра по прямым кортикоталамическим путям и на неспецифические через кортикоретикулоталамические связи. Так, например, раздражение сенсомоторной зоны коры угнетает на длительное время возникновение реакции вовлечения. Эти факты свидетельствуют о наличии между таламусом и корой больших полушарий двусторонних циклических связей, которые играют важную роль в интегративной деятельности мозга. Не случайно таламокортикальная система связана с регуляцией таких физиологически важных состояний, как смена сна и бодрствования, сохранение сознания, развитие процессов внутреннего торможения.
3.7.3 Гипоталамус
Гипоталамус — филогенетически старый отдел промежуточного мозга, который играет важную роль в поддержании постоянства внутренней среды и обеспечении интеграции функций вегетативной, эндокринной и соматической систем. Этот небольшой по объему, но важный по функциям отдел лежит на дне и по бокам третьего мозгового желудочка, вентральнее таламуса. Он включает в себя такие анатомические структуры, как серый бугор, воронку, которая заканчивается гипофизом, и мамиллярные или сосцевидные тела. Верхнюю границу гипоталамуса формируют конечная пластинка и перекрест зрительного нерва (рис. 3.13). Сбоку гипоталамус ограничен зрительным
трактом и внутренней капсулой, а сзади примыкает к среднему мозгу.
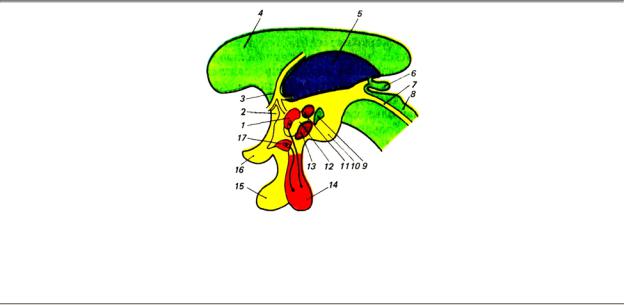
Рис. 3.13. Расположение основных ядерных групп гипоталамуса: 1 — паравентрикулярное ядро, 2 —
передняя спайка, 3 — свод, 4 — мозолистое тело, 5 — зрительный бугор (таламус), 6 — эпифиз, 7 — водопровод, 8 — средний мозг, 9 — заднее ядро, 10 — сосцевидное тело, 11 — дорсомедиальное ядро, 12 — латеральное ядро гипоталамуса, 13 — вентромедиальное ядро, 14 — нейрогипофиз, 15 — аденогипофиз, 16
—зрительный перекрест, 17 — супраоптическое ядро
Внейтральной сети гипоталамуса можно выделить несколько десятков ядер, которые топографически подразделяются на пять групп. Преоптическая группа, наиболее близкая к конечному мозгу, состоит из перивентрикулярного, медиального и латерального преоптических ядер. Передняя группа включает супрахиазматическое, супраоптическое и паравентрикулярное ядра. Отростки нейронов двух последних четко отграниченных ядер идут к задней доле гипофиза — нейрогипофизу. Средняя группа ядер гипоталамуса образована вентромедиальным и дорсомедиальным ядрами. В состав наружной группы входят латеральное гипоталамическое ядро и ядро серого бугра. И, наконец, задняя группа сформирована из заднего гипоталамического ядра, перифорникального ядра и нескольких мамиллярных ядер.
Большинство ядер гипоталамуса имеют плохо очерченные границы и, за малым исключением (супраоптическое и паравентрикулярное ядра), их трудно рассматривать как центры с узкой локализацией специфических функций. Более перспективным является деление гипоталамуса на области и зоны, обладающие определенной функциональной спецификой. Так, например, некоторые ядра преоптической и передней групп объединяются в гипофизотропную область, нейроны которой продуцируют так называемые рилизинг-факторы (либерины) и ингибирующие факторы (статины), регулирующие деятельность передней доли гипофиза — аденогипофиза.
Срединная группа ядер формирует медиальный гипоталамус, где имеются своеобразные нейроны-датчики, реагирующие на различные изменения внутренней среды организма (температуру крови, водно-электролитный состав плазмы, содержание гормонов в крови). Посредством нервных и гуморальных механизмов медиальный гипоталамус управляет деятельностью гипофиза.
Латеральный гипоталамус представляет собой безъядерную зону, в которой сосредоточены проводниковые элементы (медиальный пучок переднего мозга), формирующие пути к верхним и нижним отделам стволовой части мозга. Нервные клетки в латеральном гипоталамусе расположены диффузно. В целом гипоталамус как важный интегративный центр имеет богатые афферентные и эфферентные связи с различными отделами мозга.
Через систему волокон дорсального продольного пучка Шютца гипоталамус получает афферентный вход от ретикулярной формации среднего мозга. Параллельно этому каналу афферентные импульсы от среднего мозга могут поступать в гипоталамус по мамиллотегментальному или сосково-покрышечному тракту. Гипоталамус обладает двусторонними связями с таламусом и через передние таламические ядра с корой
больших полушарий. Афферентный вход от лимбической системы мозга осуществляется через медиальный пучок переднего мозга, проходящий в латеральном гипоталамусе.
Паллидо-гипоталамический тракт связывает гипоталамус с базальными ганглиями переднего мозга. Кроме того, афферентные входы гипоталамуса дополняются непрямыми мозжечково-гипоталамическими и вагосупраоптическими связями. Наличие последних подтверждается и электрофизиологическими данными о представительстве в гипоталамусе висцеральной афферентации, приходящей по чувствительным волокнам блуждающего и чревного нервов.
Основные эфферентные связи гипоталамуса с вегетативными и соматическими ядрами головного и спинного мозга осуществляются через полисинаптические пути ретикулярной формации ствола.
Эти пути дополняются короткими эфферентными трактами, идущими в таламус и через его посредство далее в кору больших полушарий. В системе мозгового свода проходят эфферентные волокна от гипоталамуса в лимбическую кору. Через мамиллотегментальный тракт эфферентная импульсация от гипоталамуса может достигать среднего мозга. Особо выделяется мощный пучок нисходящих волокон, идущих от супраоптического ядра гипоталамуса в нейрогипофиз.
Таким образом, сама структурная организация гипоталамуса свидетельствует о функциональной значимости этого, отдела головного мозга. Вместе с тем эта организация настолько сложна, что в вопросе об основных принципах ее построения еще остается много загадочного.
3.7.4. Роль гипоталамуса в регуляции вегетативных функций
Исследование вегетативных и соматических реакций, вызванных локальным электрическим раздражением различных областей гипоталамуса, позволило В. Гессу
(1954) выделить в этом отделе мозга две функционально дифференцированные зоны. Раздражение одной из них — задняя и латеральная области гипоталамуса — вызывает типичные симпатические эффекты: расширение зрачков, подъем кровяного давления, увеличение частоты сердечных сокращений, прекращение перистальтики кишечника и т. д. Разрушение данной зоны, напротив, приводило к длительному снижению тонуса симпатической нервной системы и контрастному изменению всех перечисленных выше показателей. Гесс назвал область заднего гипоталамуса эрготропной и допустил, что здесь локализованы высшие центры симпатической нервной системы.
Другая зона, охватывающая преоптическую и переднюю области гипоталамуса,
получила название трофотропной, так как при ее раздражении наблюдались все признаки общего возбуждения парасимпатической нервной системы, сопровождавшиеся реакциями, направленными на восстановление и сохранение резервов организма.
Однако дальнейшие исследования показали, что гипотеза о наличии двух антагонистических зон гипоталамуса является слишком обобщенной и не может объяснить различные функции этого отдела мозга. Так были получены факты, противоречащие узкой локализации симпатических и парасимпатических центров в гипоталамусе. Оказалось, что сосудосуживающий эффект может быть получен при раздражении и задней, и передней областей гипоталамуса и, следовательно, симпатические нейроны присутствуют в этих обеих областях. Равным образом клетки, которые активируют постганглионарные холинэргические сосудорасширяющие нейроны, тоже довольно широко распространены в пределах гипоталамуса (см. разд. 5.5.3).
Детальное исследование изменений в сердечно-сосудистой системе показало, что локальное раздражение определенных отделов гипоталамуса сопровождается диаметрально противоположными изменениями кровотока в различных органах. Например, при увеличении кровотока в скелетных мышцах может наблюдаться его снижение в сосудах кожи и органах брюшной полости. При раздражении гипоталамуса