

4.2. Генетика поведения инфузорий
Ресничные одноклеточные Paramecium и Tetrahymena представляют собой прекрасные модели для изучения генетики поведение простейших. При изучении этих объектов можно использовать и поведенческий, и электрофизиологический, и биохимический и генетический подход. Анализ поведения простейших, как правило, включает изучение физиологического состояния клетки, поиск поведенческих мутантов и их отбор, а также исследование влияния различных химических веществ (лекарственных препаратов и пр.).
Поскольку парамеции являются довольно крупными (до 250 мкм в диаметре), они легко могут быть использованы для наблюдения поведения, поисков поведенческих мутантов и электрофизиологического анализа. Tetrahymena обладает сравнительно высокой скоростью гомологической рекомбинации и используется для создания трансгенных линий и стабильных нокаутных мутантов по отдельным генам.
Поведение простейших особенно подробно изучено на примере Paramecium aurelia. Локомоторные реакции парамеции находятся под контролем поверхностной мембраны. Направление и частота биения ресничек коррелируют со сдвигом электрического потенциала мембраны, обусловленным изменениями потенциалзависимой проводимости кальция. Изменения направления движения ресничек вызывают изменение направления перемещения парамеции.
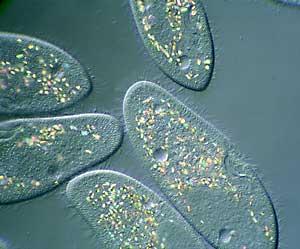
Рис. 4.2. Paramecium aurelia.
Репертуар поведения свободноживущих инфузорий заключается в том, что при действии химического или физического стимула они демонстрируют смену направления биения ресничек. Показано, что подобные локомоторные реакции находятся под контролем поверхностной мембраны. Направление и частота биения ресничек коррелируют со сдвигом электрического потенциала мембраны, который обусловлен изменениями потенциалзависимой проводимости кальция. Изменения направления движения ресничек вызывают изменение направления перемещения парамеции.
Роль специфических ионных каналов в каскадной передаче сигналов и в последующих изменениях подвижности инфузорий была выявлена при сравнении клеток дикого типа с некоторыми поведенческими мутантами.
Ученые получили сотни мутантных линий Paramecium aurelia. При помощи поведенческого, электрофизиологического и генетического анализов в этих работах показаны физико-химические механизмы адаптивного поведения простейших.
Установлено, что большинство мутаций, которые затрагивают поведение, локализованы в несцепленных локусах и наследуются согласно менделеевским законам. Электрофизиологический анализ показал, что следствием подобных мутаций зачастую является нарушение или исчезновение одного или более ионных токов, деполяризующих или реполяризующих мембрану при реакции избегания.
Известны мутанты Pawn (что значит пешка, передвижение которой по шахматной доске имеет определенные ограничения). Они в отличие от особей дикого типа не могут плыть назад и могут быть температурочувствительными. Всего выделяют несколько несцепленных локусов, мутанты по которым характеризуются фенотипом Pawn: pwA, pwB, pwC.
Ещё одна мутация Paramecium tetraurelia (d4-662), сначала характеризовавшаяся как четвёртый локус Pawn – мутация pwD – фактически оказалась аллелью pwB (pwB662). Показано также, что аллели pwB взаимодействуют между собой.
Мутация Pawn проявляется в специфическом моногенном дефекте, связанном с отсутствием возбудимости мембраны, несущей реснички. Реснички при этом не затрагиваются мутацией. При морфологических исследованиях у парамеций были обнаружены нити, связывающие основания ресничек (базальные тельца). Предполагалось, что система этих нитей координирует движение ресничек, однако доказательств этому не получено.
Прочтение генома инфузории Tetrahymena thermophila показало, что генетический код, используемый инфузорией, оказался несколько иным, чем у всех других организмов: стоп-кодоны у тетрахимены могут использоваться для кодирования аминокислот. Кроме того, выяснилось, что эта инфузория умеет эффективно удалять из рабочих копий своего генома весь «мусор» — повторяющиеся и чужеродные фрагменты. Потомство, однако, всегда получает в наследство «неочищенный» вариант генома.
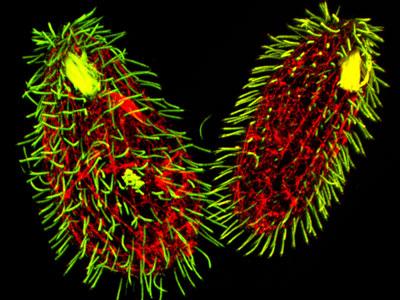
Рис. 4.3. Tetrahymena thermophila
У тетрахимены, как у всех инфузорий, есть два клеточных ядра — маленький микронуклеус, содержащий две копии генома (пять пар хромосом), и большой макронуклеус с многократно дуплицированным геномом (несколько сотен хромосом).
Микронуклеус хранит наследственный материал для передачи следующим поколениям, но его гены практически не работают. Гены макронуклеуса, наоборот, активно работают - транскрибируются — считываются и используются для синтеза белков, но они не могут быть переданы потомкам.
Инфузории, как и другие простейшие, размножаются делением, причем делятся оба ядра. Однако такое бесполое размножение не может длиться бесконечно долго: рано или поздно инфузории должны сливаться попарно и обмениваться наследственным материалом. В ходе этого полового процесса (конъюгации) макронуклеус разрушается вместе со всеми своими генами.
В обмене наследственным материалом участвуют только микронуклеусы. Поэтому следующее поколение инфузорий (образовавшееся после полового процесса) получает лишь те гены, которые хранились в микронуклеусах родителей. Микронуклеус снова делится пополам, и одно из дочерних ядер начинает расти и превращается в новый макронуклеус.
Ученые установили, что в ходе образования макронуклеуса геном не только многократно дублируется, но и модифицируется: длинные хромосомы разрезаются на несколько более мелких, некоторые фрагменты генома удаляются, другие меняются местами.
Исследователям удалось прочесть геном макронуклеуса. Он оказался неожиданно велик, причем не столько по своей длине (105 миллионов пар оснований — это в 30 раз меньше, чем у человека, и примерно во столько же раз больше, чем у среднестатистической бактерии), сколько по числу генов. Геном одноклеточной тетрахимены содержит свыше 27 тысяч генов, которые кодирую различные белки. Это примерно столько же, сколько геном человека и значительно больше, чем другие прочтенные геномы одноклеточных.
Еще одна необычная особенность генома инфузории состоит в чрезвычайно низком содержании повторяющихся нуклеотидных последовательностей и мобильных генетических элементов (транспозонов): они занимают всего 2% генома (для сравнения - у человека — около 50%).
Функциональная роль повторяющихся последовательностей остается неясной. Возможно, некоторые из этих последовательностей важны для регуляции экспрессии генов. Кроме того, известно, что многочисленные повторы и мобильные элементы придают геному пластичность и повышают вероятность различных геномных перестроек (например, транслокаций — перемещений фрагментов ДНК на другую хромосому или на новое место в той же хромосоме).
Исследователи пришли к выводу, что низкое содержание повторов и мобильных элементов в геноме макронуклеуса тетрахимены объясняется тем, что почти весь «мусор», имеющийся в геноме микронуклеуса, удаляется при формировании большого ядра.
Повторы, очевидно, не нужны для нормальной работы генов и для жизнеобеспечения организма, они удаляются из «рабочей копии» генома. Но они передаются потомкам, возможно для того, чтобы геном сохранял пластичность. Вероятно, это адаптация, направленная на оптимизацию эволюционного процесса.
В геноме инфузории используется только один стоп-кодон, а не три, как у большинства других живых организмов. У тетрахимены роль стоп-кодона выполняет только одна последовательность нуклеотидов — УГА. Два другие классические стоп-кодона (УАА и УАГ) у тетрахимены кодируют аминокислоту глутамин.
Впрочем, и триплет УГА у тетрахимены не всегда обозначает конец гена. В некоторых случаях (это зависит от нуклеотидного «контекста») эти три нуклеотида кодируют редкую аминокислоту селеноцистеин.
Тетрахимена оказалась единственным из изученных на сегодняшний день организмов, у которого могут транслироваться все 64 триплета.
Ученые получили поведенческие мутанты Tetrahymena, неспособные плыть назад вследствие специфического нокаута гена, кодирующего динеин.
У Tetrahymena thermophila дикого типа при помощи видео дифференциальной интерференционной контрастной микроскопии обнаружена интересная поведенческая реакция, способствующая завершению цитокинеза. Этот процесс, названный ротокинезом облегчает физическое разделение дочерних клеток.
Ротокинез включает постоянное однонаправленное вращение одной из образующихся дочерних клеток вокруг оси, проходящей вдоль цитоплазматического мостика, соединяющего дочерние клетки. Предполагают, что вращающий момент, генерируемый ротокинезом, может ослаблять цитоплазматический мостик между предполагаемыми дочерними клетками, тем самым увеличивая вероятность успешного цитокинеза. Механизм, который обеспечивает именно такое финальное разделение клеток в конце цитокинеза изучен не полностью.
Нокаутные по генам, кодирующим kinesin-II гомологичные молекулярные моторы Kin1p и Kin2p, линии Tetrahymena thermophila парализованы вследствие полной утраты ресничек и подвержены частым нарушениям цитокинеза. Наблюдения за живыми делящимися клетками показали, что формирование борозды деления у двойных kinesin-II нокаутов идёт нормально вплоть до последней стадии разделения клеток.
У тетрахимены описана также мутация dcc (defective in ciliogenesis and cytokinesis – дефект биогенеза ресничек и цитокинеза), вследствие которой у особей наблюдаются нарушения биогенеза ресничек. Они неспособны восстанавливать подвижность после децилиации и неспособны к полноценному цитокинезу.
Преимущества Paramecium и Tetrahymena формируют основу для генетического анализа функциональных компонентов сенсорной трансдукции и адаптационных реакций этих животных.