

6.8. Метод локализации фокуса действия мутации на карте презумптивных органов дрозофилы.
Очень часто мутация, проявляющаяся на уровне поведения дрозофилы, затрагивает соответствующие рецепторные или эффекторные системы. Однако такая ситуация наблюдается не всегда. Ученые изучили гиперкинетические мутанты у дрозофилы. Поведение этих мутантов характеризуется тикообразными подергиваниями лап во время анестезии. Это эффект обнаруживается у гомозиготных по этой мутации самок и у самцов.
При изучении различных мозаиков по этой мутации обнаружили, что, для проявления описанного поведенческого дефекта, грудной сегмент должен быть мутантным. Генотип головы, ноги, брюшка не влияет на гиперкинетическое поведение. Иными словами, мутантная по генотипу нога очень часто не проявляет описанного синдрома. У билатеральных мозаиков поведение левых ног не зависит от поведения правых.
Данные, полученные при изучении различных мозаиков, продемонстрировали высокую корреляцию между внешним фенотипом кутикулы груди и проявлением тика. Однако эта корреляция не была абсолютной, у некоторых особей с мутантной грудью не наблюдалось подергивания лап и, наоборот, особи с нормальным генотипом груди иногда проявляли патологическое поведение. Эти данные можно истолковать следующим образом: не всегда генотип кутикулы груди отражает генотип структуры, ответственной за данный синдром. В некоторых, хотя и редких, случаях генотип морфологических маркеров не соответствует генотипу регулируемой изучаемое поведение структуры. Поэтому, естественно, встает вопрос, как локализовать те внутренние структуры, которые ответственны за формирование мутантного фенотипа, и как идентифицировать их генотип у мозаиков.
Для этого был предложен метод эмбрионального картирования или метод локализации мозаичных границ на бластуле. В основу метода был положен разработанный Стёртевантом принцип картирования генов на хромосоме путем определения частоты рекомбинаций, которая является функцией расстояний между ними.
В 1929 г. Стёртевант предложил тот же принцип для составления мозаичной карты бластодермы. Частота появления особей с разным генотипом тех или иных частей тела должна зависеть от расстояния между участками бластодермы, из которых возникают эти части. Стёртевант изучил около 400 мозаиков, но не обработал полученные данные. Через 40 лет по его данным была составлена карта бластулы («fate map»).
С. Бензер также с целью составления карты бластулы изучил 700 мозаичных мух. Он оценил расстояния между эмбриональным положением различных структур и построил карту расположения эмбриональных зачатков внешних частей тела на поверхности бластулы (рис. 6.6.).
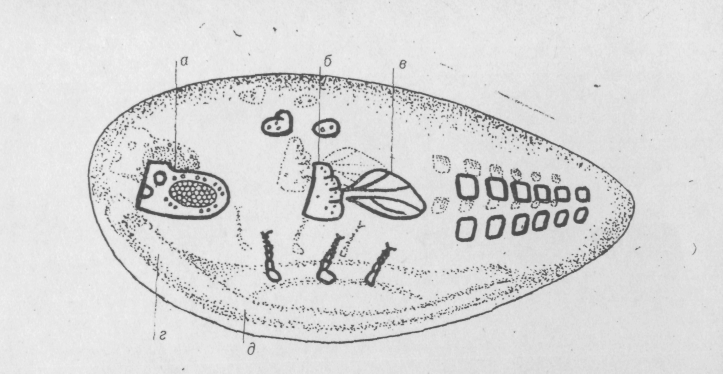
Рис.6.6. Карта эмбриональных зачатков на поверхности бластулы.
а— голова; б — грудь; в — крыло; г — головной мозг;
д — вентральная нервная система.
Бензер ввел единицу расстояния между эмбриональным положением различных структур, названную в память Стёртеванта Стёртом. Один Стёрт — это расстояние, эквивалентное возможности, что в 1% случаев среди всех изученных мозаичных мух две структуры будут иметь различный генотип.