

59.Глаз, глазное яблоко и вспомогательные органы. Мышцы глазного яблока, их иннервация.
Периферический отдел зрительной сенсорной системы представлен рецепторами, расположенными в сетчатке глаза. Но прежде чем изучать строение сетчатки, рассмотрим устройство самого глазного яблока.
Внешний вид глаза. Глазное яблоко расположено в глазнице черепа. У детей оно имеет шаровидную форму, у взрослых его переднезадний размер несколько превышает поперечный и вертикальный и составляет примерно 24 мм. Различают переднийи задний полюсы глаза. Линия, соединяющая оба полюса глазного яблока, называется его осью. Зрительный нерв входит в глазное яблоко несколько медиальнее его заднего полюса.
Глазное яблоко окружено тремя оболочками: наружной – фиброзной, средней – сосудистой и внутренней – сетчатой (см. Атл.). В центре глазного яблока находится ядро, которое состоит из хрусталика, стекловидного тела и водянистой влаги – это преломляющие среды глаза. Впереди хрусталика лежит передняя камера глаза, также заполненная жидкостью.
Оболочки глазного яблока. Фиброзная оболочка (tunica fibrosa bulbi) наружная и самая прочная, благодаря ей глазное яблоко поддерживает свою форму. Она представлена двумя отделами. Передний, занимающий 1/5 ее поверхности, образует прозрачная, сильно вогнутая роговица, обладающая светопреломляющим свойством; задний – белочная оболочка – склера, по цвету напоминающая белок вареного куриного яйца.
Роговица (cornea) состоит в основном из плотной соединительной ткани (прозрачное собственное вещество роговицы). Спереди она покрыта многослойным плоским неороговевающим эпителием, а сзади, со стороны наружной камеры глаза, выстлана однослойным эпителием – эндотелием. Раздражение нервных окончаний, которыми пронизан наружный эпителий роговицы, вызывает рефлекторное моргание и слезотечение. Кровеносные сосуды в роговице отсутствуют.
Склера (sclera) покрывает задний, больший отдел глазного яблока. Она также образована плотной соединительной тканью, но не прозрачна из-за большого количества коллагеновых и эластических волокон и несколько иного состава межклеточного вещества. В передней части склера переходит в роговицу. Границей между ними является тонкий полупрозрачный ободок –лимб (край) роговицы. На границе между роговицей и склерой проходит венозный синус, по которому из глаза оттекает венозная кровь и лимфа. Эпителий роговицы переходит здесь вконъюнктиву, выстилающую переднюю часть белочной оболочки. В задней части глаза в области выхода волокон зрительного нерва в склере образуются многочисленные отверстия (решетчатая пластинка). По ее краям склера наиболее массивна и переходит в соединительнотканную оболочку нерва. Утолщение склеры наблюдается также впереди экватора глазного яблока, где к ней прикрепляются четыре прямые мышцы глаза. Кровеносные сосуды проходят через склеру к сосудистой оболочке и цилиарному телу.
Сосудистая оболочка (tunica vasculosa bulbi) состоит из трех различных по структуре и функциям частей: собственно сосудистой оболочки, ресничного тела и радужки.
Собственно сосудистая оболочка (chorioidea) рыхло соединяется со склерой. Между ними располагаются лимфатические щели. Оболочка тонкая (до 0,2 мм), состоит из трех слоев (пластинок). Самый наружный слой – надсосудистая пластинка – образован эндотелием, эластическими волокнами, соединенными со склерой, между которыми расположены многочисленные пигментные клетки и контактирующие с ними нервные волокна. Сосудистая пластинка занимает среднюю часть оболочки. В ней располагаются крупные сосуды, преимущественно вены, между которыми лежат соединительнотканные волокна и пигментные клетки. В глубоком слое сосудистой оболочки – хориокапиллярной пластинке –залегают крупные капилляры синусоидного типа. Их сеть особенно хорошо развита в области желтого пятна сетчатки (см. Атл.). Строение капилляров таково, что кровь быстро переходит из артериального русла в венозное. На границе с сетчаткой лежит полупроницаемая базальная мембрана (стекловидная перегородка, мембрана Бруха), которая содержит эластические волокна.
На экваторе сосудистая оболочка прободается четырьмя венами, выходящими на равном расстоянии друг от друга (см. Атл.). В переднем отделе она без резких границ переходит в ресничное тело.
Ресничное тело (corpus ciliare), имеющее вид валика, вдается внутрь глазного яблока там, где белочная оболочка переходит в роговицу (см. Атл.). Задний край тела переходит в собственно сосудистую оболочку, а от переднего отходит до 70ресничных отростков. От них берут начало упругие тонкие волоконца, другим своим концом прикрепляющиеся к капсуле хрусталика по его экватору. Эти волоконца образуют поддерживающий хрусталик аппарат, или ресничный поясок (циннову связку). Внутри его, между волоконцами, остается пространство, окружающее хрусталик по экватору и содержащее водянистую влагу. В соединительнотканной основе ресничного тела, кроме сосудов, содержатся гладкие мышечные волокна, меридиональные, радиальные и циркулярные, составляющие ресничную мышцу, обеспечивающую аккомодацию.
Радужная оболочка, или радужка (iris), имеет вид диска с отверстием посередине – зрачком и находится позади прозрачной роговицы. Своим наружным краем радужка переходит в ресничное тело, а внутренним, свободным, ограничивает – зрачок. В ее соединительнотканной основе заложены сосуды, пигмент и гладкая мускулатура. От количества и глубины залегания пигмента зависит цвет глаз, варьирующий от светло-голубого до черного. Красноватый оттенок глаз альбиносов, совершенно лишенных пигмента, вызван просвечиванием кровеносных сосудов. Мышечные волокна радужки имеют двойное направление. По радиусам расположены волокна мышцы, расширяющей зрачок, вокруг зрачкового края радужки находятся круговые волокна мышцы, суживающей зрачок. Эти мышцы придают радужке значение диафрагмы, регулирующей поступление света в глаз.
Сетчатая оболочка, или сетчатка (retina), является внутренней оболочкой глазного яблока. Ее наружная поверхность прилежит к сосудистой оболочке, а внутренняя – к стекловидному телу. В сетчатке различают три части, из которых задняя, большая – зрительная часть – светочувствительная, в ней расположены рецепторные клетки. На уровне задней границы ресничного тела она переходит в ресничную часть в виде неровной линии –зубчатой каймы. Передняя часть сетчатки – радужинная – подстилает радужку. Последние две части нечувствительны к свету.
Зрительная часть сетчатки имеет сложное микроскопическое строение, она состоит из 10 слоев (см. Атл.). Самым наружным слоем, прилегающим к сосудистой оболочке, служит пигментный эпителий. Непосредственно за ним располагается слой нейроэпителия, содержащий рецепторные клетки. Эти клетки из-за формы своих наружных сегментов получили название палочеки колбочек. Их периферические отростки, образующие второй слой сетчатки, вдаются в слой пигментного эпителия. Число рецепторов в глазу человека огромно (палочек около 130 млн., колбочек – 6– 7 млн). Колбочки – рецепторы "цвета", они преобладают в средней части сетчатки; палочки, обеспечивающие сумеречное зрение и располагаются в ее боковых частях. Центральные отростки зрительных рецепторных клеток приходят в соприкосновение с биполярными игоризонтальными клетками, которые, в свою очередь, контактируют с ганглиозными клетками. Нейриты последних образуют зрительный нерв. В слое рецепторных клеток отсутствуют кровеносные сосуды, питательные вещества поступают сюда из хориокапиллярной пластинки сосудистой оболочки.
Таким образом, в сетчатке рецепторные клетки находятся в самом наружном слое. Световой поток проходит через стекловидное тело и попадает на глубокие слои сетчатки. Для того, чтобы достичь палочек и колбочек свет должен пройти через всю толщину сетчатки до пигментного слоя.
Количественная оценка клеточного состава сетчатки показала, что численность клеток в различных ее слоях неодинакова. Она убывает в ряду рецепторные клетки – биполярные клетки – ганглиозные клетки. Это свидетельствует о том, что на одной биполярной клетке суммируются афферентные импульсы от нескольких фоторецепторных клеток, а на одной ганглиозной клетке – от нескольких биполярных. Наряду с этим, в слое биполярных клеток присутствуют горизонтальные клетки, которые образуют синаптические контакты с рецепторными и биполярными клетками, а в слое ганглиозных клеток –амакриновые клетки, контактирующие с би-полярами и ганглиозными клетками.
Все описанные клетки сетчатки, кроме пигментных, развиваются из стенки мозгового пузыря, т.е. аналогичны нейронам мозга. Кроме них в сетчатке развиваются и глиальные клетки, которые получили название радиальных (Мюллеровых) клеток. Это длинные узкие клетки, ядро которых располагается примерно на уровне ядер биполярных клеток. Радиальные глиальные клетки контактируют с палочками и колбочками и имеют в этой части сетчатки большое скопление филаментозного вещества. Ранее его считали мембраной и называли наружной пограничной мембраной. Микроворсинки на апикальной части глиальных клеток проникают между рецепторными клетками.
В задней части сетчатки выделяются два участка – диск и желтое пятно. Диск – место выхода из глазного яблока зрительного нерва; здесь сетчатка не содержит светочувствительных элементов. В области диска в сетчатку вступает питающая ее артерия и выходит вена. Оба сосуда проходят внутри зрительного нерва. Желтое пятно находится почти точно на заднем полюсе глаза, это самое чувствительное к свету место сетчатки, так как здесь сосредоточено большое количество колбочек. Середина пятна углубляется вцентральную ямку. Линию, соединяющую середину переднего полюса глаза с центральной ямкой, называют оптической осью глаза. Для лучшего видения глаз устанавливается так, чтобы рассматриваемый предмет и центральная ямка находились на одной оси.
Волокна зрительного нерва покрываются миелиновой оболочкой только после прохождения через решетчатую пластинку. Диаметр нерва при этом увеличивается.
Ядро глазного яблока. Хрусталик (lens) – плотное тело в виде двояковыпуклого зерна чечевицы (см. Атл.). Закругленный его край называют экватором. Хрусталик лишен сосудов и нервов, совершенно прозрачен и покрыт бесструктурной прозрачнойкапсулой. Задняя поверхность хрусталика вдается в стекловидное тело, расположенное позади него, а передняя прилежит к радужке. Хрусталик укреплен ресничным пояском. При сокращении мышечных волокон ресничного тела натяжение пояска ослабевает и хрусталик, не испытывая ограничивающего давления своей капсулы, становится более выпуклым. Это повышает его преломляющую способность. Изменение кривизны хрусталика обусловливает приспособление глаза к ясному видению разноудаленных предметов и называетсяаккомодацией.
Хрусталик – наиболее мощная преломляющая среда глаза (показатель преломления – 1,43). С возрастом он уплотняется и уплощается, и аккомодация ослабевает.
Стекловидное тело (corpus vitreum) заполняет в глазу все пространство между сетчаткой сзади и хрусталиком спереди. Оно плотно прилегает к сетчатке, способствуя прилеганию пигментного и наружного ее слоев и содействует фиксации хрусталика. Стекловидное тело состоит из прозрачного студенистого межклеточного вещества и не имеет сосудов. Преломляющая способность его равна 1,33.
Водянистая влага выделяется кровеносными сосудами ресничных отростков и радужки. Она заполняет полости:переднюю камеру глаза, расположенную между роговицей и радужкой, и заднюю камеру – между радужкой и хрусталиком с его пояском. Обе эти камеры сообщаются через зрачок, и водянистая влага омывает радужку, частично ресничное тело и хрусталик. Водянистая влага очень слабо преломляет свет. Ее отток осуществляется через венозный синус.
Вспомогательные аппараты глаза. К вспомогательным аппаратам органа зрения относятся веки, слезная железа, мышцы глазного яблока, жировое тело орбиты и фасция (см. Атл.).
Верхнее и нижнее веки, ограничивая глазную щель и дополняя спереди глазницу, образуют подвижную защиту глазного яблока. Основой век служит полулунная пластинка плотной волокнистой соединительной ткани, пронизанная своеобразно измененными сальными железами. Последние открываются на свободном крае век и выделяют жирный беловатый секрет. Снаружи от пластинки расположена вековая часть круговой мышцы глаза, относящейся к мимическим. У свободного края век расположены влагалища корневых луковиц ресниц. Внутренняя поверхность век выстлана оболочкой –конъюнктивой, которая продолжается на глазном яблоке, покрывая его свободную поверхность. Конъюнктивой ограничивается конъюнктивальный мешок, который содержит слезную жидкость, омывающую свободную поверхность глаза и обладающую бактерицидным свойством
У внутреннего угла глаза между краями век образуется пространство – слезное озеро, на дне его лежит маленькое возвышение – слезное мясцо. На крае обоих век в этом месте находится по небольшому отверстию – слезной точке, это начало слезного канальца. Латеральнее слезного мясца конъюнктива образует вертикально поставленную складочку – рудимент мигательной перепонки глаза низших позвоночных.
Слезная железа расположена в верхнелатеральной части глазницы, в одноименной ямке лобной кости. Выводные протоки железы (в числе 10–12) открываются в латеральной части конъюнктивального мешка Слезная жидкость предохраняет роговицу от высыхания и смывает с нее пылевые частицы Из конъюнктивального мешка слезная жидкость частично испаряется, а частично оттекает через слезные канальцы.Начинаясь слезными точками у медиального угла глаза, канальцы направляются под кожей век к слезному мешку,расположенному на медиальной стенке глазницы, и впадают в него. Слезный мешок, суживаясь книзу, переходит в слезно-носовой проток, который помещается в одноименном костном канале и открывается в нижний носовой ход
Глазное яблоко приводится в движение шестью глазными мышцами: четырьмя прямыми и двумя косыми
Прямые мышцы глаза берут начало от фиброзного кольца в окружности зрительного канала черепа Мышцы прикрепляются к глазному яблоку впереди его экватора по четырем сторонам – снаружи, изнутри, сверху и снизу, соответственно чему они и называются – латеральная, медиальная, верхняя и нижняя. Эти мышцы благодаря своему положению вращают глазное яблоко вокруг вертикальной и фронтальной осей. Но только латеральная и медиальная мышцы поворачивают глаз прямо наружу и внутрь; верхняя же и нижняя – не только вверх и вниз, а немного и внутрь.
Верхняя косая мышца тоже начинается от упомянутого выше фиброзного кольца, направляется к медиальному углу глазницы и перекидывается здесь через блок лобной кости. После этого мышца меняет направление и под острым углом подходит к верхнелатеральной стороне глазного яблока, позади экватора, где и прикрепляется. При сокращении мышца поворачивает глазное яблоко так, что зрачок обращается вниз и наружу.
Нижняя косая мышца начинается от глазничной поверхности верхнечелюстной кости, идет поперек глазницы, огибая снизу глазное яблоко, и прикрепляется к его наружной поверхности позади экватора. Мышца направляет зрачок вверх и наружу, вращая глаз, как и верхняя косая, по сагиттальной оси.
Жировое тело заполняет пространство между стенками глазницы и глазным яблоком с его мышцами. Жировое тело образует мягкую и эластичную обкладку глазного яблока.
Фасция отделяет жировое тело от глазного яблока; между ними остается щелевидное пространство, которое обеспечивает подвижность глазного яблока.
Проводниковый и центральный отделы зрительной сенсорной системы. Проводниковый отдел начинается всетчатке (рис. 3.62). Нейриты ее ганглиозных клеток складываются в зрительные нервы, которые, войдя через зрительные каналы в полость черепа, образуют перекрест. У низших позвоночных (амфибий, рептилий) в перекресте участвуют все волокна зрительных нервов, поэтому движения левого и правого глаза независимы, поля их зрения разделены, зрение монокулярное. У обезьян и человека перекрещивается около половины волокон зрительных нервов. Это обеспечивает согласованные движения глазных яблок и бинокулярное зрение. После перекреста каждый нерв называется зрительным путем (трактом). В составе каждого зрительного тракта идут волокна от латеральной половины сетчатки глаза своей стороны и медиальной – другого глаза. Тракт огибает ножку мозга и разделяется на два корешка. Один из них заканчивается
сохраняется только у клеток, лежащих на экваторе. Дочерние клетки перемещаются в глубину хрусталика, перестают делиться, в них начинается синтез кристаллинов. Затем клетки утрачивают ядра и органеллы и остаются в виде волокон. Волокна, расположенные в центре хрусталика, в его ядре образовались еще во внутриутробный период. Ближе к периферии, в коре хрусталика, лежат волокна, образовавшиеся в более поздний период.
При погружении хрусталика под эктодерму, последняя наползает на него и образует радужную оболочку и ресничное тело. Вокруг закладки глаза из мезенхимы развиваются сосудистая оболочка и склера. К склере прикрепляются мышечные волокна, которые затем превращаются в мышцы глазного яблока. Эпителий, покрывающий
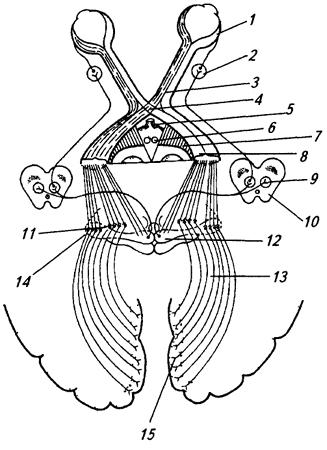 Рис.
3.62. Схема зрительных путей:
Рис.
3.62. Схема зрительных путей:
1 – сетчатка; 2 – ресничный узел; 3 – зрительный нерв; 4 – перекрест зрительных нервов (хиазма); 5 – гипоталамус; 6 – мамиллярные тела; 7 – ножка мозга; 8 – зрительный тракт; 9 – ядро глазодвигательного нерва; 10 – средний мозг; 11 – подушка таламуса; 12 – верхние бугорки четверохолмия; 13 – зрительная лучистость; 14 – латеральное коленчатое тело; 15 – зрительная область коры
в верхнем двухолмии. Его волокна идут к ниже расположенным эффекторным ядрам ствола (глазодвигательного и других нервов), а также к мотонейронам спинного мозга (тектоспинальный путь). Благодаря этому осуществляются рефлекторные ответы на зрительные раздражения (например, непроизвольные движения головы и глаз). Нейроны верхнего двухолмия участвуют в рецепции движущихся объектов. На поверхности двухолмия имеется упорядоченная проекция сетчатки (ретинотопия). В глубоких слоях серого вещества лежат мотонейроны, отвечающие за движение глаз в определенном направлении. Оба типа нейронов (чувствительные поверхностные и моторные глубокие) связаны между собой. Нейроны, занимающие центральное положение, получают проекции от слуховой системы и соматической чувствительности верхней половины тела (голова, верхние конечности).
Другой корешок направляется к подушке зрительного буграи латеральному коленчатому телу. Нейроны латерального коленчатого тела у приматов сгруппированы в 6 слоев (рис. 3.63). К каждому слою подходят аксоны от сетчатки только одного глаза. Кроме того, каждый участок сетчатки проецируется на определенную группу нейронов. Наибольшее количество нейронов воспринимает информацию от центральной части сетчатки, в том числе желтого пятна. Таким образом, здесь проявляется топическая организация проекции различных зон сетчатки. Нейроны латерального коленчатого тела обладают также цветовой чувствительностью. В подушке и латеральном коленчатом теле зрительные импульсы переключаются на следующий нейрон, волокна которого в составе зрительной лучистости идут к коре затылочной области больших полушарий.
Центральным проекционным полем зрительной системы является поле 17.
закладку снаружи, становится роговицей. В ее внутренний слой мигрируют эндотелиальные клетки из сосудистой оболочки.
На 7 неделе эмбрионального периода начинают развиваться веки. Они возникают в виде складок кожи над роговицей, соединяются друг с другом и срастаются. На 6–7 месяце пренатального развития веки раскрываются. Слезные железы появляются на 9 неделе.
Положение глаз на голове также меняется в процессе развития. На 6 неделе глаза расположены по бокам головы, затем, по мере роста лицевых структур, глаза перемещаются на переднюю поверхность лица и к 10 неделе располагаются на ней почти как у взрослого.
От него ассоциативные волокна идут к полям 18 и 19, где расположены соответственно вторичная и третичная зрительные проекции. В эти поля проецируется также часть волокон зрительной лучистости. Поле 17 связано также с полями 21 и 7, а, кроме того, с верхним двухолмием, претектальной областью, подушкой таламуса и латеральным коленчатым телом, а поле 19 – с полями 17, 18, 21 и 7. Из поля 18 эфферентные волокна идут к претектальной области и подушке таламуса. Волокна, имеющие разную толщину и идущие от разных подкорковых структур, оканчиваются в разных слоях коры. Здесь также прослеживается четкая топическая организация; центральная часть сетчатки имеет более обширную проекцию. Электрофизиологически было установлено, что 84% нейронов зрительной коры реагируют при одновременной стимуляции сетчаток двух глаз, т.е. они ответственны за бинокулярное зрение (рис 3.64). Нейроны зрительной коры различаются не только по способности реагировать на моно- или бинокулярное раздражение, но и по реакции на движение объекта, его размер и т.д., нейроны коры, образующие вертикальные связи друг с другом, складываются в колонкуСоседние колонки, реагирующие на возбуждение одного или другого глаза, были названы колонками глазодоминантности (рис. 3.64). Такие колонки были продемонстрированы морфологически: введение в один глаз меченых аминокислот позволило выявить на горизонтальных (тангенциальных) срезах коры причудливо пересекающиеся полосы (рис. 3.64, Б, В). Темные полосы на этой фотографии соответствуют локализации меченой аминокислоты (ЗН-пролин), введенной в один глаз. Аминокислота была доставлена из сетчатки в нейроны коры аксоплазматическим транспортом по отросткам нейронов зрительных путей. Аналогичным образом была продемонстрирована слоистая организация латерального коленчатого тела (рис. 3.63).
В зрительном восприятии принимает участие нижневисочная область коры (поле 21). С ней связывают дифференциацию предметов по форме, отношение их к определенной категории, установлению равнозначности объектов, которые проецируются в разные зоны сетчатки. На активность нейронов этой зоны оказывают влияние миндалина и гиппокамп. Поле 7 принимает участие в организации пространственного зрения.
Для полного анализа предмета (его величины, расстояния от глаз и т.д.) к ощущениям от раздражения сетчатки прибавляются ощущения от раздражения проприорецепторов аккомодационных мышц ресничного тела, мышц, суживающих и расширяющих зрачок.
60.Анатомия органа слуха. Внутренне строение улитки: спиральный (кортиев) орган.
см лекцию
61.Анатомия вестибулярного аппарата.
Вестибулярный аппарат — орган равновесия позволяющий определять положение и перемещение тела в пространстве. Вестибулярный аппарат расположен в полости внутреннего уха. Он состоит из двух перепончатых мешочков — круглого и овального — и трех полукружных каналов. Мешочки заполнены эндолимфой и выстланы изнутри чувствительными рецепторными клетками. Рецепторы расположены на возвышениях — макулах. Ворсинками рецепторные клетки пронизывают желеобразную мембрану.
Мембрана содержит кристаллики карбоната натрия — отолиты. Возбуждение рецепторов органа равновесия происходит вследствие скольжения отолитовой мембраны по волоскам и их сгибания. Отолитовый прибор сигнализирует главным образом о положении головы, а также об ускорении или замедлении прямолинейного движения. Своей тяжестью отолиты раздражают оказавшиеся под ними рецепторы, сигналы которых указывают направление сил гравитации.
Полукружные каналы представляют собой изогнутые полукругом трубки, открытые обоими концами в овальный мешочек и расположенные в трех взаимно перпендикулярных плоскостях. На одном из своих концов каждый полукружный канал образует вздутие — ампулу. В ампуле находится рецепторный орган. Каналы расположены таким образом, что любое движение головы находит точное отражение в сигналах от полукружных каналов. Сигналы рецепторов полукружных каналов сообщают главным образом о движениях головы в трех взаимно перпендикулярных направлениях. Вестибулярные центры тесно связаны с мозжечком и гипоталамусом. Поэтому при укачивании у человека теряется координация движений, возникает тошнота.
62.Орган обоняния, вкуса: строение и проводящие пути.
Вкусовая и обонятельная сенсорные системы позволяют человеку оценивать химический состав пищи и окружающего воздуха. По этой причине их объединяют под названием хемосенсорные системы. Сюда же относятся внутренностные хеморецепторы (каротидного синуса, пищеварительного тракта и другие). Химическая рецепция – одна из наиболее филогенетически древних форм связи организма со средой.
Рецепторный отдел вкусовой сенсорной системы расположен в ротовой полости и представлен вкусовыми рецепторными клетками. Они собраны во вкусовые почки, которые находятся
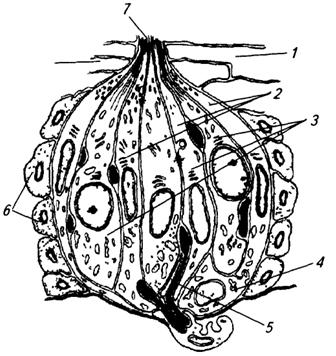 Рис.
3.73. Строение вкусовой почки:
Рис.
3.73. Строение вкусовой почки:
1 – эпителиальные клетки; 2 – опорные клетки; 3 – рецепторные клетки; 4 –базальная клетка; 5 – нервные волокна; 6 – клетки окружающего эпителия; 7 – вкусовая пора
главным образом в сосочках на дорсальной поверхности языка – грибовидных, листовидных и желобовидных. Одиночные вкусовые почки рассеяны в слизистой оболочке мягкого неба, миндалин, задней стенки глотки и надгортанника. У детей область их распространения шире, чем у взрослых; к старости их количество уменьшается.
Наиболее типичное строение у человека имеют вкусовые почки желобовидных сосочков (рис. 3.73). Каждая почка представляет собой овальное образование, занимающее всю толщину эпителия и открывающееся на его поверхность вкусовой порой. Почка имеет около 70 мкм в высоту, 40 мкм в диаметре и образована 40–60 удлиненными клетками, располагающимися наподобие долек в апельсине. Среди клеток вкусовых почек различают рецепторные, опорные и базальные. Первые два вида клеток занимают всю длину почки от ее базальной части до вкусовой поры. Относительно рецепторной функции этих клеток до сих пор идут споры. Предполагается, что опорные клетки также могут участвовать в рецепторном процессе.
Вкусовые рецепторные клетки являются вторичночувствующими. В их апикальную мембрану, обращенную к вкусовой поре, встроены рецепторные молекулы, с которыми связываются различные химические вещества. В результате мембрана клетки переходит в возбужденное состояние. Через синаптические контакты в базолатеральной части клетки возбуждение передается на нервное волокно, а далее в головной мозг. Человек различает четыре основных вкуса (сладкий, соленый, горький, кислый) и несколько дополнительных (металлический, щелочной и т.д.). Рецепция вкусовых веществ становится возможной тогда, когда эти вещества попадают на поверхность языка, растворяются в слюне, проходят через вкусовую пору и достигают апикальной мембраны рецепторных клеток.
Срок жизни рецепторных и опорных клеток невелик – около 10 дней. Их обновление происходит за счет митотического деления клеток в базальной части почки.
Проводниковый и центральный отделы вкусовой сенсорной системы. Вкусовые афферентные волокна от передних двух третей языка, от вкусовых почек грибовидных сосочков передней части языка и нескольких листовидных сосочков, проходят в составе лицевого нерва (барабанная струна – chorda tympani) (ветвь VII пары), а от задней трети, задних листовидных и желобовидных – в составе языкоглоточного нерва (IX пара) (рис. 3.74). Вкусовые почки задней стенки ротовой полости и глотки иннервируются блуждающим нервом (X пара). Эти волокна являются периферическими отростками нейронов, лежащих в ганглиях этих нервов: VII пара – в коленчатом ганглии, IX пара – в каменистом ганглии. Волокна всех нервов, по которым передается вкусовая чувствительность, оканчиваются в ядре одиночного пути (nucleus tractus solitarius). Отсюда восходящие волокна следуют к нейронам дорсальной части моста (парабрахиальное ядро) и к вентральным ядрам таламуса (рис. 3.75). От таламуса часть импульсов идет в новую кору – в нижнюю часть постцентральной извилины (поле 43).
 Рис.
3.75. Проводящие пути вкусовой сенсорной
системы:
Рис.
3.75. Проводящие пути вкусовой сенсорной
системы:
1 – соматосенсорная область языка (прикосновение, температура); 2 –таламус; 3 – вкусовое ядро моста; 4 – ядро одиночного пути; 5 – латеральный гипоталамус; 6 – островковая область коры (вкус, осязание, температура); 7 – соматосенсорная вкусовая область (тактильная, температурная чувствительность); ВА – висцеральные афференты; VII, IX, X – черепно-мозговые нервы
Предполагают, что с помощью этой проекции происходит различение вкуса. Другая часть волокон от таламуса направляется в структуры лимбической системы (парагиппокампову извилину, гиппокамп, миндалину и гипоталамус). Эти структуры обеспечивают мотивационную окраску вкусовых ощущений, участие в ней процессов памяти, которые лежат в основе приобретаемых с возрастом вкусовых предпочтений.
В слизистой оболочке передней части языка оканчиваются также волокна тройничного нерва (V пара). Они попадают сюда в составе язычного нерва (рис. 3.74). По этим волокнам передается тактильная, температурная, болевая и другая чувствительность с поверхности языка, которая дополняет информацию о свойствах стимула в ротовой полости.
С помощью обоняния человек способен различать тысячи запахов, но тем не менее он относится к микросматикам, так как у человека эта система значительно менее развита, чем у животных, которые с ее помощью ориентируются в окружающей среде.
Периферическим отделом обонятельной сенсорной системы являются рецепторные клетки в эпителиальной (обонятельной) выстилке носовой полости. Она расположена в верхней носовой раковине и соответствующей части носовой перегородки, отличается желтоватым цветом (из-за присутствия пигмента в клетках) и занимает в носовой полости около 2,5–5 см2.
Слизистая оболочка носовой полости в области обонятельной выстилки несколько утолщена по сравнению с остальной слизистой. Она образована рецепторными и опорными клетками (см. Атл.). Обонятельные рецепторные клетки представляют собой первичночувствующие клетки. В их апикальной части расположен длинный тонкий дендрит, заканчивающийся булавовидным утолщением. От утолщения отходят многочисленные реснички, имеющие обычное строение и погруженные в слизь. Эту слизь выделяют опорные клетки и железы, лежащие под эпителиальным пластом (Боуменовы железы). В базальной части клетки расположен длинный аксон. Немиелинизированные аксоны многих рецепторных клеток образуют под эпителием довольно толстые пучки, называемые обонятельными волокнами (fila olfactoria). Эти аксоны проходят в отверстия продырявленной пластинки решетчатой кости и направляются к обонятельной луковице, лежащей на нижней поверхности головного мозга (см. рис. 3.15). Возбуждение рецепторных клеток происходит при взаимодействии стимула с ресничками, затем оно по аксону передается в мозг. Хотя обонятельные клетки являются нейронами, они, в отличие от последних, способны к обновлению. Срок жизни этих клеток составляет примерно 60 дней, после чего они дегенерируют и фагоцитируются. Замена рецепторных клеток происходит за счет деления базальных клеток обонятельной выстилки.
Проводниковый и центральный отделы обонятельной сенсорной системы. В обонятельной луковице различают пять слоев, расположенных концентрически (рис. 3.72):
1 слой образуют волокна обонятельного нерва – отростки обонятельных рецепторных клеток;
262
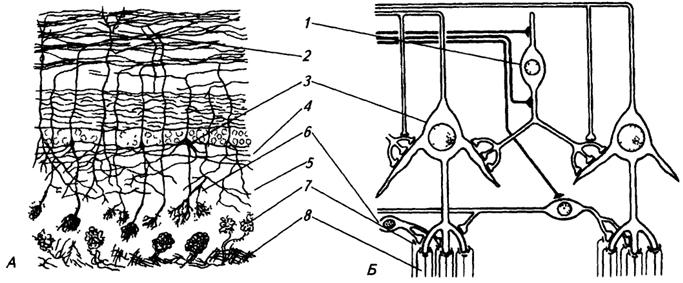 Рис.
3.72. Обонятельная луковица:
Рис.
3.72. Обонятельная луковица:
А – рисунок с гистологического препарата; Б – схема, 1 – клетки зерна; 2 – зернистый слой;3 – митральные клетки, 4 – внутренний и 5 – наружный сетевидные слои; 6 – перигломерулярные клетки, 7 – гломерулы; 8 – отростки обонятельных рецепторных клеток
2 слой образован гломерулами диаметром 100–200 мкм, здесь происходит синаптический контакт обонятельных волокон с отростками нейронов следующего порядка,
3 слой – наружный сетевидный (плексиформный), образован перигломерулярными клетками, контактирующими с несколькими гломерулами каждая,
4 слой – внутренний сетевидный (плексиформный), содержит самые крупные клетки обонятельной луковицы –митральные клетки (второй нейрон). Это крупные нейроны, апикальные дендриты которых образуют по одной гломеруле во 2 слое, а аксоны формируют обонятельный тракт. В пределах луковицы аксоны митральных клеток образуют коллатерали, контактирующие с другими клетками. Во время электрофизиологических экспериментов было установлено, что запаховая стимуляция вызывает разную активность митральных клеток. Клетки, расположенные в различных участках обонятельной луковицы, реагируют на определенные виды запахов;
5 слой – зернистый, образуют клетки-зерна, на которых оканчиваются эфферентные волокна, приходящие из центра. Эти клетки способны контролировать активность митральных клеток.
От обонятельной луковицы отходит обонятельный тракт,образованный аксонами митральных клеток. По нему обонятельные сигналы поступают в другие области мозга (см. Атл.). Тракт оканчивается латеральной и медиальной обонятельными полосками. Через латеральную обонятельную полоску импульсы попадают главным образом в древнюю коруобонятельного треугольника, где лежит третий нейрон, а затем в миндалину. Волокна медиальной обонятельной полоскизаканчиваются в старой коре подмозолистого поля, прозрачной перегородке, в клетках серого вещества в глубине борозды мозолистого тела. Обогнув последнее, они доходят до гиппокампа. Здесь берут начало волокна свода – проекционной системы старой коры, заканчивающегося частично в прозрачной перегородке и в мамиллярном теле гипоталамуса. От него начинаются мамилло-таламический путь, идущий к одному из ядер (переднему) таламуса, и мамилло-тектальный путь,оканчивающийся в межножковом ядре покрышки ножек мозга, откуда импульсы проводятся в другие эфферентные ядра центральной нервной системы. Из переднего ядра таламуса импульсы направляются в кору лимбической области. Кроме того, из первичной обонятельной коры нервные волокна доходят до медиовентрального ядра таламуса, где имеются также входы от вкусовой системы. Аксоны нейронов этого ядра идут к фронтальной (лобной) области коры, которую рассматривают как высший интегративный центр обонятельной системы. Гипоталамус, гиппокамп, миндалевидное тело и лимбическая область коры взаимосвязаны, они входят в состав лимбической системы и принимают участие в формировании эмоциональных реакций, а также в регулировании деятельности внутренних органов. Связь обонятельных путей с этими структурами объясняет участие обоняния в питании, эмоциональном статусе и т.д.
В крыше каждой носовой области дифференцируется обонятельная область. Обонятельные рецепторные клетки – биполярные нейроны – дифференцируются в самом эпителии среди длинных столбчатых клеток, называемых опорными клетками. Отростки рецепторных клеток, обращенные к поверхности эпителия, образуют расширения – булавы, увенчанные пучком видоизмененных ресничек, которые несут на своей поверхности рецепторы химических веществ. Противоположные отростки этих клеток удлиняются и устанавливают связь с нейронами в обонятельной луковице, которые передают нервные импульсы в соответствующие центры головного мозга.