

Приобретенные пороки седца
.pdf11
верстия (valvula foraminis ovalis). Левое ушко более узкое и длинное, чем правое,
оно отграничено от предсердия хорошо выраженным перехватом. Левый желудо-
чек имеет коническую форму. Его емкость от 130 до 220 мл, толщина стенки 11— 14 мм. Масса левого желудочка 130—150 г. Из-за закругленности левого края сердца передняя и задняя стенки левого желудочка нерезко разграничены, меди-
альная стенка соответствует межжелудочковой перегородке. Ближайший к отвер-
стию аорты участок левого желудочка называется артериальным конусом. Внут-
ренняя поверхность желудочка, за исключением перегородки, имеет многочис-
ленные мясистые трабекулы. Вверху располагаются два отверстия: слева и спере-
ди — овальное левое предсердно-желудочковое (его продольный размер состав-
ляет 23—37 мм, поперечный — 17—33 мм), справа и сзади — отверстие аорты.
Клапан левого предсердно-желудочкового отверстия (митральный) имеет чаще всего две створки и соответственно две сосочковые мышцы — переднюю и зад-
нюю. Клапан аорты образован тремя полулунными заслонками — задней, правой и левой. Начальная часть аорты в месте расположения клапана расширена (диа-
метр ее достигает 22—30 мм) и имеет три углубления — синусы аорты. Стенки сердца образуются тремя оболочками: наружной — эпикардом, внутренней — эн-
докардом и расположенной между ними мышечной оболочкой — миокардом.
Эпикард — висцеральная пластинка пери-
карда — является серозной оболочкой.
Рис. 8. Микропрепарат нормального миокарда:
видны параллельно расположенные мышечные во-
локна; окраска гематоксилином и эозином; ×180.
Он состоит из тонкой пластинки соедини-
тельной ткани с различным расположением эластических и коллагеновых волокон, по-
крытой с поверхности мезотелием. Миокард (рис. 8) составляет основную массу
12
стенки сердца. Миокард желудочков отделен от миокарда предсердий фиброзны-
ми кольцами, от которых начинаются пучки волокон миокарда. В миокарде желу-
дочков условно можно выделить наружный, средний и внутренний (глубокий)
слои. Наружные слои миокарда желудочков общие. Ход волокон наружного и внутреннего слоев имеет вид редкой спирали; средний слой пучков миокарда циркулярный. Гистологически ткань миокарда отличается от поперечнополосатой скелетной мышечной ткани рядом признаков, в т.ч. меньшими размерами клеток миокарда (кардиомиоцитов) и саркомер, наличием в клетке одного ядра, соедине-
нием кардиомиоцитов последовательно друг с другом по типу конец в конец по-
средством вставочных дисков и др. Около 30—40% объема кардиомиоцита зани-
мают митохондрии. Особая насыщенность кардиомиоцитов митохондриями от-
ражает высокий уровень метаболизма ткани, обладающей непрерывной активно-
стью. В миокарде имеется особая система волокон, обладающих способностью проводить импульсы ко всем мышечным слоям сердца и координировать после-
довательность сокращения стенки его камер.
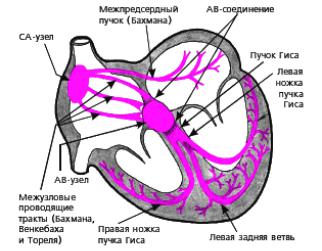
Рис. 9. Проводящая система сердца.
Эти специализированные мышечные волокна составляют проводящую систе-
му сердца. Она состоит из синусно-
предсердного и предсердно-желудочко-
вого узлов и пучков (предсердных, меж-
узловых соединительных, предсердно-желудочкового и его ветвей и др.) - рис. 9.
В ткани проводящей системы сердца, более адаптированной к анаэробному мета-
болизму, чем сократительный миокард, митохондрии занимают около 10% кле-
точного объема, а миофибриллы — около 20%. Эндокард выстилает полость сердца, включая сосочковые мышцы, сухожильные хорды, трабекулы и клапаны.
13
В желудочках эндокард тоньше, чем в предсердиях. Он, как и эпикард, состоит из двух слоев: субэндотелиального и коллагеново-эластического, покрытых эндоте-
лием. Створка клапана сердца представляет собой складку эндокарда, в которой имеется соединительнотканная прослойка. Иннервация сердца происходит из сердечного сплетения, расположенного под эпикардом, большей частью в стенках предсердий, меньшей — в стенках желудочков (рис. 4). Оно образовано ветвями грудного аортального сплетения, а также имеет сердечные ганглии, содержащие синапсы пре - и постганглионарных парасимпатических нервных волокон. В со-
ставе ветвей грудного аортального сплетения к сердцу подходят постганглионар-
ные симпатические, преганглионарные парасимпатические и чувствительные нервные волокна. Волокна сердечного сплетения формируют вторичные интра-
муральные сплетения с чувствительными и мигательными волокнами. Крово-
снабжение сердца осуществляется обычно правой и левой венечной артериями,
отходящими от луковицы аорты (рис. 4). В зависимости от преобладающего зна-
чения какой-либо из них в обеспечении сердца кровью различают правовенечный
(рис. 6) и левовенечный (рис. 7), а также равномерный типы кровоснабжения Ле-
вая венечная артерия делится на огибающую и переднюю межжелудочковую вет-
ви. От огибающей артерии отходят несколько ветвей, в т.ч. анастомотическая пе-
редняя, предсердно-желудочковые, левая краевая, промежуточная предсердная,
задняя левого желудочка, а также ветви синусно-предсердного и предсердно-
желудочкового узлов и предсердные ветви. От передней межжелудочковой арте-
рии отделяются ветви артериального конуса, латеральная и перегородочные меж-
желудочковые. Правая венечная артерия отдает ветвь артериального конуса, вет-
ви синусно-предсердного и предсердно-желудочкового узлов, предсердные и промежуточные предсердные ветви, правую краевую, заднюю межжелудочковую
(от нее отходят перегородочные межжелудочковые ветви) и правую заднелате-
ральную ветвь. Артерии сердца ветвятся во всех его оболочках. Благодаря ана-
стомозам в сердце может быть коллатеральное кровообращение. Отток крови из вен его стенки происходит в основном в венечный синус, впадающий в правое

14
предсердие. Кроме того, кровь оттекает непосредственно в правое предсердие че-
рез передние вены сердца. Лимфоотток осуществляется из лимфокапиллярной се-
ти эндокарда в сосуды миокарда, а из сетей миокарда и эпикарда — в субэпикар-
диальные лимфатические сосуды. Из них формируются правый и левый главные лимфатические сосуды, впадающие в лимфатические узлы средостения.
ФИЗИОЛОГИЯ
Сердце работает как насос, нагнетая кровь в артерии в период сокращения желудочков, или систолу, и заполняясь кровью из вен в период расслабления, или диастолу. Деятельность сердца как насоса является основным источником механической энергии дви-
жения крови в малом и большом кругах кровооб-
ращения, благодаря чему поддерживается непре-
рывность обмена веществ и энергии в организме
(рис. 10).
Рис. 10. Схема кровообращения. 1 и 2 - правый и левый желудочки; 3 - малый круг кровообращения; 4 - система церебральных сосудов; 5 - кровоснабжение желудка, 6 - се-
лезенки, 7 - кишечника; 8 - большой круг кровообращения;
9 - кровоснабжение почек; 10 - воротная вена; 11 - система кровоснабжения печени
Энергетическое обеспечение этой деятельности зависит почти исключительно от выработки АТФ в окислительном фосфорилиро-
вании, протекающем в митохондриях и требующем наличия молекулярного кис-
лорода (прекращение поступления кислорода приводит к быстрому уменьшению сократительной способности миокарда). Для аэробного метаболизма сердца глав-
ными энергетическими субстратами являются жирные кислоты, глюкоза, лактат,
пируват и кетоновые тела, поступающие в кардиомиоциты из плазмы крови, и в
15
меньшей степени — аминокислоты. При наличии жирных кислот и углеводов преимущественными энергетическими субстратами для сердца являются жирные кислоты, на окисление которых используется около 70% кислорода, потребляемо-
го сердцем. Транспорт глюкозы в кардиомиоциты увеличивается при повышении концентрации глюкозы, в присутствии инсулина, катехоламинов, а также в усло-
виях гипоксии, и уменьшается при повышении концентрации жирных кислот. Из плазмы крови в кардиомиоциты проникают только свободные жирные кислоты
(СЖК). Липопротеины и триглицериды плазмы крови могут использоваться в энергетическом обмене сердца только после их расщепления до СЖК фермента-
ми. В клетках СЖК активируются с использованием АТФ и образованием ацетил-
КоА. Продукт окисления СЖК ацетил-КоА, подвергается окислению в цикле три-
карбоновых кислот до углекислоты и воды. Общая скорость аэробного окисления СЖК и углеводов связана со скоростью потребления кислорода, которая связана линейной зависимостью с работой, выполняемой сердцем. При изменении крово-
обращения от состояния, соответствующего физическому покою организма, до состояния при максимальных физических нагрузках, скорость потребления кис-
лорода сердцем может меняться в пределах от 50 до 300 мкг атомов на 1 г сухого веса ткани в 1 мин. Содержание в сердце высокоэргических фосфатов в этих условиях практически не меняется, т.к. их количество, потраченное на сокраще-
ние сердца, быстро восполняется за счет синтеза в митохондриях. При этом важ-
ное значение имеет не только образование АТФ из АДФ, но и транспорт энергии в кардиомиоцитах, т.к. для сокращения сердца используется только АТФ, локали-
зованная в миофибриллах и около мембран. Транспортную функцию осуществля-
ет фосфокреатин (ФКр) с участием креатинкиназы, которая катализирует реакцию АТФ + креатин ФКр + АДФ, поддерживая тем самым соотношение АТФ: АДФ на постоянном уровне. Преобразование энергии высокоэргических фосфатов в механическую работу сердечных сокращений связано со специфическими физио-
логическими функциями отдельных структур сердца, способствующих преобра-
зованию химической энергии в механическую в определенном ритме. Соответ-

16
ственно, кроме свойства сократимости важными объектами физиологического ис-
следования являются такие свойства сердца, как автоматизм, возбудимость (спо-
собность возбуждаться под действием раздражителя), проводимость, рефрактер-
ность и др. Импульсы возбуждения периодически возникают в сердце под влия-
нием процессов, протекающих в нем самом. Это явление получило название ав-
томатизма. Способностью к автоматизму обладает специфическая мышечная ткань, формирующая синусно-предсердный узел и проводящую систему сердца.
Рис. 11. Трансмембранный потенциал действия.
АРП и ОРП - абсолютный и относительный ре-
фрактерный периоды
На мембранах клеток специфической му-
скулатуры сердца зарождаются электри-
ческие импульсы, переходящие на рабочий миокард и вызывающие его сокраще-
ния. Эти процессы связаны с деполяризацией клеточных мембран, которые в со-
стоянии покоя клетки всегда поляризованы вследствие разной концентрации Nа+
и К+ на поверхности и внутри клетки из-за неодинаковой проницаемости мембра-
ны для этих ионов (рис.12). В состоянии покоя мембрана кардиомиоцита почти непроницаема для Nа+ и частично проницаема для К+, который под действием процесса диффузии выходит из клетки, увеличивая на поверхности мембраны по-
ложительный заряд. При этом внутренняя поверхность мембраны приобретает от-
рицательный заряд, образуется потенциал покоя мембраны порядка 60—80 мВ.
Возбуждение клетки связано с увеличением проницаемости мембраны для Nа+,
поступление которого в клетку сопровождается деполяризацией мембраны и ре-
версией потенциала на ее поверхности, т.е. наружная поверхность мембраны при-
обретает отрицательный электрический заряд. При этом формируется потенциал действия, превышающий на своем пике величину потенциала покоя, достигая значений до 100 мВ и более. Потенциал действия деполяризует мембраны сосед-
них клеток, в результате чего они генерируют собственные потенциалы действия
17
— происходит распространение процесса возбуждения по клеткам миокарда. В
отличие от волокон сократительного миокарда, мембрана клеток, способных к ав-
томатизму, не имеет после выхода из возбуждения постоянного потенциала по-
коя, т.к. сохраняет некоторую степень проницаемости для Nа+. Вследствие пере-
мещения этих ионов внутрь клетки и одновременного снижения проницаемости для К+ возникает постепенное уменьшение положительного заряда на поверхно-
сти мембраны — развивается так называемая медленная диастолическая деполя-
ризация. Когда уровень потенциала покоя уменьшится по сравнению с исходным приблизительно на 20 мВ, возникает резкое увеличение проницаемости мембраны для Nа+, в результате чего Nа+ лавинообразно поступает внутрь клетки, вызывая деполяризацию мембраны и формируя потенциал действия. Участок, в котором автоматически зарождаются импульсы, ведущие к сокращению сердца называют водителем ритма, или Пейсмекером. В нормальных условиях им является синус-
но-предсердный узел (рис.9). Особенностью проводящей системы предсердий и желудочков является способность почти каждой из ее клеток (кроме клеток пред-
сердно-желудочкового узла) самостоятельно генерировать импульсы возбужде-
ния, т. е. она, как и синусно-предсердный узел, обладает автоматизмом. Суще-
ствует так называемый градиент автоматизма, выражающийся в убывании часто-
ты генерации импульсов клетками проводящей системы по мере удаления их от синусно-предсердного узла. Клетки синусно-предсердного узла человека в покое спонтанно генерируют ритмические импульсы возбуждения частотой 60—80 им-
пульсов в минуту, клетки пучка Гиса и его ножек — частотой 30—40 импульсов в минуту, а волокна Пуркинье — частотой около 20 импульсов в минуту. В обыч-
ных условиях автоматизм всех участков проводящей системы подавляется часты-
ми импульсами, поступающими к ним из синусно-предсердного узла, но в случае поражения последнего, водителем ритма может стать ниже расположенный отдел проводящей системы.

18
Рис.12. Рас-
простране-
ние возбуж-
дения по предсерди-
ям. а -
начальное возбуждение правого предсердия, б - возбуждение правого и левого предсердий, в -
конечное возбуждение левого предсердия. Р1, Р2 и Р3 - моментные векторы деполяризации
Рис.13. Рас-
пространение
возбуждения по сократи-
тельному миокарду же-
лудочков. а - деполяризация межжелудочковой перегородки (0,02 с), б - деполяризация вер-
хушки, передней, задней и боковой стенок желудочков (0,04 - 0,05 с), в - деполяризация базаль-
ных отделов желудочков и межжелудочковой перегородки (0,06 - 0,08 с)
Возникнув в синусно-предсердном узле, возбуждение по специальным внут-
рипредсердным проводящим путям, а также диффузно, распространяется по мио-
карду предсердий (рис.12 а,б,в,) достигает предсердно-желудочкового узла и по-
сле некоторой задержки в нем (благодаря которой кровь во время сокращения предсердий заполняет полость еще расслабленных желудочков) распространяется по пучку Гиса и волокнам Пуркинье — к волокнам сократительного миокарда
(рис. 13 а,б,в). В миокарде предсердий и желудочков скорость проведения воз-
буждения составляет 0,9—1,0 м/сек, в волокнах предсердно-желудочкового узла
— 0,05 м/сек, пучке Гиса — 1—1,5 м/сек, волокнах Пуркинье — 3 м/сек. Быстрое проведение через волокна Пуркинье обеспечивает почти одновременное возбуж-
дение различных участков миокарда желудочков, что повышает мощность сокра-
щения сердца и эффективность работы желудочков по нагнетанию крови (рис.13).
Время охвата поверхности желудочков возбуждением составляет 10—15 м/сек.
Электрические потенциалы, возникновение которых связано с распространением
19
возбуждения по сердцу, можно зарегистрировать при помощи электродов, нало-
женных на поверхность тела. Потенциалы действия мембран сердечных волокон представляют собой пусковой механизм, включающий серию внутриклеточных процессов, сопрягающих возбуждение с сокращением миофибрилл. Сокращение мышцы происходит без изменения длины актиновых и миозиновых нитей сарко-
мера — основной сократительной единицы мышечной ткани. Укорочение мы-
шечного волокна достигается за счет вдвигания актиновых нитей между миозино-
выми благодаря веслообразным движениям поперечных мостиков — выступаю-
щих участков миозиновой нити, образованных из меромиозина, обладающего АТФ-азной активностью. При расслаблении мышцы актиновые нити отодвигают-
ся назад и занимают прежнее положение по отношению к нитям миозина. Акти-
новые нити состоят из цепочек молекул белка актина, на поверхности которых находятся тонкие нити из белка тропомиозина, блокирующего центры взаимодей-
ствия актина с миозином. Тропомиозин образует комплекс с белком тропонином,
обладающим высоким сродством к Са2+. Процесс сокращения миокарда запуска-
ется Са2+, который поступает к сократительным белкам из цистерн саркоплазма-
тического ретикулума под воздействием импульса возбуждения. Са2+ связывает-
ся тропонином, что вызывает изменение пространственного расположения тропо-
нин-тропомиозинового комплекса на актиновой нити, снимая тормозное действие его на активные центры актина. В результате происходит ассоциация актина с миозином — образование актомиозина, идентифицируемое с сокращением, и
расщепление АТФ, освобождающее энергию для скольжения актиновых нитей.
Процесс расслабления миокарда возникает в результате удаления ионов кальция от тропонина под действием реполяризации мембраны и связывания их сарко-
плазматическим ретикулумом, а также вследствие «откачивания» ионов кальция насосами клеточных мембран в межклеточную жидкость. При повторных элек-
трических раздражениях остановленного сердца, происходит постепенное повы-
шение концентрации ионов кальция внутри клетки, вследствие чего сила каждого последующего сокращения постепенно возрастает до тех пор, пока сокращения не

20
достигнув максимальной величины. Это постепенное возрастание силы сокраще-
ний получило название «лестница Боудича». Возможность появления сокращений в ответ на раздражение сердца электрическим током используется в современных методах нормализации ритма сердца с помощью портативных электростимулято-
ров. Сердце нагнетает кровь в сосудистую систему благодаря периодическому по-
следовательному сокращению мышечных клеток предсердий и желудочков.
Рис.14. Схема движения крови в сердце (направление пока-
зано стрелками). 1-v.cava superior; 2- truncus brachiocephalicus; 3- a.carotis communis sinistra; 4- a.subclavia sinistra; 5 - pars descendens aortae; 6 – a.pulmonalis sinistra; 7- atrium sinistra; 8 – w.pulmonalis sinistri; 9- ventriculus sinistrer; 10ven- triculus dexter; 11 – v.cava inferior; 12 – atrium dextrum; 13truncus pulmonalis; 14- a. pulmonalis dextra
Внутри сердца, благодаря функции клапанов, кровь движется только в одном направлении: в фазе диастолы — из предсердий в желу-
дочки, в фазе систолы желудочков — из правого желудочка в легочный ствол, из левого — в аорту (рис.14). Захлопывание и открытие клапанов, связаны с измене-
нием направления градиента давления между сообщающимися камерами в фазы систолы и диастолы желудочков. Движение клапанов и перемещение крови вме-
сте с изменением напряжения стенок сердца, сопровождаются звуковыми фено-
менами, в частности образованием тонов сердца. Около 2/3 объема крови, посту-
пающей в желудочки в фазу диастолы, притекает в связи с положительным дав-
лением крови в экстракардиальных венах, и 1/3 подкачивается в желудочки в фазу систолы предсердий. Предсердия являются резервуаром для притекающей крови,
легко меняющим свою емкость благодаря небольшой толщине их стенок и нали-
чию добавочных емкостей — ушек предсердий, способных при расправлении вместить значительный объем крови. При каждом сокращении сердца, правый и