

Александров Ю.И., Анохин К.В. - Нейрон. Обработка сигналов. Пластичность. Моделирование_ Фундаментальное руководство (2008)
.pdfЭто достигается путем продукции клетками глии в зоне повреждения целого ряда факторов, ингибирующих рост аксонов и вызывающих коллапс конуса роста: свободных радикалов, окиси азота, производных арахидоновой кислоты, белков N1-35 и N1-250 (Николлс, 2003). Интересно, что поврежденные пути в ЦНС могут все-таки регенерировать, если в очаг поражения имплантировать суспензии ШК или же сегмент периферического нерва с оболочками из ШК (например, седалищного нерва). Так, при имплантации сегмента седалищного нерва в виде «мостика» между перерезанными участками спинного мозга мыши или крысы нарушенные волокна регенерировали и прорастали вдоль этого мостика в зону повреждения (Николлс, 2003). Экспериментально показано, что такие мостики, эффективно способствующие восстановлению функций мозга, могут организовываться между разными участками ЦНС, вклю- чая кору (David, Aguayo, 1981; Николлс, 2003) (рис. 14.13).
Ðèñ. 14.13. Использование «глиальных мостиков» для восстановления поврежденных нервных связей (по: David, Aguayo, 1981). (А) Место размещения имплантанта (куска седалищного нерва), соединяющего (в виде «мостика») разобщенные в результате перерезки области спинного (spinal cord) и продолговатого (medulla) мозга. (В) Области нейронов (зачернены), аксоны которых прорастали через «глиальный мостик». Видно, что эти участки располагались вблизи нижнего (6.5 mm) и верхнего (4 mm) концов имплантанта.
14.3.2. Роль макроглии в миелинизации аксонов
Бесспорна и наиболее хорошо изучена роль глии в формировании и регенерации миелиновых оболочек, которые изолируют аксоны от окру-
450
жающей среды и за счет своего высокого сопротивления обеспечивают надежность и высокую скорость проведения возбуждения (рис. 14.14, 14.15).
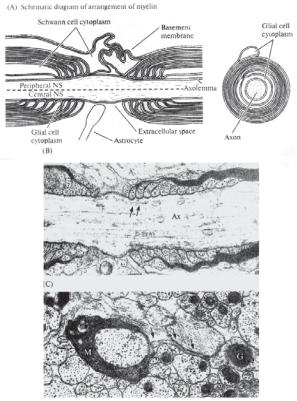
Ðèñ. 14.14. Олигодендроглия (в ЦНС, Central NS) и Шванновские клетки (в периферической нервной системе, Peripheral NS) формируют миелиновые оболочки аксонов (по: Николлс с соавт., 2003). (А) Расположение глии в районе перехватов Ранвье (слева) и поперечный срез миелинизированного аксона (справа). Обозначения: Schwann cell cytoplasm — цитоплазма Шванновской клетки; Glial cell cytoplasm — цитоплазма олигодендроцита; Astrocyte — отросток астроцита; Extracellular space — экстраклеточное пространство; Basement membrane — базальная мембрана; Axon — аксон; Axolemma — аксоплазма.
(B) Вид перехвата Ранвье волокна ЦНС крысы под электронным микроскопом. Обозначения: Ax — аксон; стрелками обозначены складки миелиновой оболочки. (С) Электронная микрофотография поперечного среза миелинизированного волокна в области перехвата Ранвье. Обозначения: М — миелин, G — астроцит, отросток которого (указан стрелками) подходит к перехвату Ранвье.
451
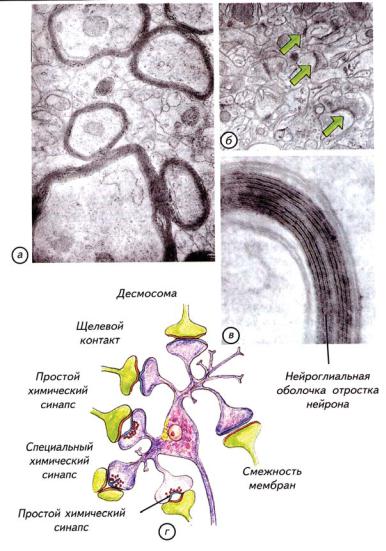
Ðèñ. 14.15. Электронные микрофотографии оболочек аксонов нервных клеток (а, в; поперечные срезы отростков), образованные Шванновскими клетками; (г) — схема основных типов межнейронных синапсов; (б) — вид синапти- ческих контактов под электронным микроскопом (обозначены зелеными стрелками) (по: Савельев, 2005).
452
Миелинизацию и ремиелинизацию (при повреждениях) аксонов нейронов в ЦНС осуществляют олигодендроциты, а в периферической нервной системе — Шванновские клетки.
Дефицит миелинизации в нервной системе (наследственный или приобретенный) является причиной ряда неврологических заболеваний, делающих человека инвалидом (рассеянный склероз, паралич, слепота и др.). Поэтому актуальной проблемой современных исследований в этой области является изучение молекулярных механизмов динамического взаимодействия глии и нейронов в процессах миелинизации, димиелинизации и ремиелинизации. К настоящему времени идентифицированы основные белки, обеспечивающие это взаимодействие. В частности, выявлена критически важная роль в формировании периферического миелина белка РМР22, выделяемого Шванновскими клетками (Notterpek et al., 1997). Точечная мутация этого белка, выражающаяся в замене всего лишь одной аминокислоты лейцина на пролин, является причиной наследственной нейропатии у человека. Обнаружено, что аденозин, образующийся из АТФ в результате реакции дефосфорилизации, стимулирует рост олигодендроглии и выработку миелина (Stevens et al., 2002).
Если олигодендроциты и Шванновские клетки обеспечивают миелинизацию аксонов, то другие клетки макроглии — астроциты — уча- ствуют при этом в кластеризации натриевых каналов в перехватах Ранвье (Rasband et al., 1999). В частности, выдвинута и обсуждается гипотеза о том, что молекулярные комплексы, образующие натриевые каналы, переносятся от астроцитов и встраиваются в мембраны перехватов Ранвье (Shrager et al., 1985). В пользу такого предположения свидетельствуют данные о высокой плотности натриевых каналов в мембранах астроцитов (Ritchie et al., 1990).
14.3.3. Роль глии в снабжении нейронов питательными веществами и регуляции кровоснабжения мозга
Гипотеза о «метаболических функциях» глии была сформулирована в 1883 г. знаменитым итальянским гистологом, лауреатом Нобелевской премии по физиологии и медицине 1906 г. (совместно с Рамон-и- Кахалом) К. Гольджи. Гипотеза базировалась на морфологических данных об интенсивных ветвлениях концевых отростков глии вокруг сосудистых капилляров мозга. Современные данные подтверждают возможность участия глии в снабжении нейронов питательными веще-
453
ствами (Ransom, Fern, 1997). В частности, показано, что Мюллеровские клетки сетчатки вырабатывают и выделяют в экстраклеточное пространство молочную кислоту (лактат), потребность в которой у клеток сет- чатки резко увеличивается в состоянии повышенной активности или же недостатке кислорода.
Имеются косвенные данные об участии астроцитов в снабжении нейронов кислородом и глюкозой путем локальной регуляции просвета кровеносных капилляров. Выдвинутая Paulson & Newman (1987) гипотеза о механизмах такой регуляции, по сути, не отличается от «метаболической гипотезы» К. Гольджи, но предполагает обратный характер взаимодействия глии и нейронов в процессе снабжения последних питательными веществами. А именно: не глия переносит метаболиты от крови к нейронам (гипотеза Гольджи), а сами нейроны путем локальной активации глии вызывают расширение сосудов и, как следствие, локально увеличивают приток глюкозы и кислорода (гипотеза Paulson
&Newman, 1987).
14.3.4.Роль глии в поддержании гомеостаза ионного состава экстраклеточной среды
Высокая чувствительность глии к ионам калия послужила основанием для выдвижения гипотезы о том, что глиальные клетки вклю- чены в процесс перераспределения ионов калия во внеклеточной среде, образуя в нервной системе своеобразный «пространственный буфер экстраклеточной концентрации калия» (Николлс и др., 2003). Необходимость в таком перераспределении ионов калия возникает в связи с неравномерной активностью нейронов в разных локусах нервной сети: в областях повышенной активности экстраклеточная концентрация калия выше, нежели в других отделах. Этот дисбаланс в пространственном распределении ионов калия является результатом активации потенциалзависимых калиевых каналов, выход наружу ионов калия через которые обеспечивает реполяризацию мембраны активированных нейронов и формирует задний фронт потенциала действия. Таким образом, активность нейрона, сопровождаемая нарастанием выходящего калиевого тока, приводит к накоплению экстраклеточного калия и, как следствие, к увеличению входящего калиевого тока (деполяризации) в рядом расположенной клетке глии. Далее входящий калиевый ток из локуса повышенной концентрации калия распространяется по электрохимическому градиенту в другие части
454
глиальной клетки (с более низкой концентрацией калия) и через электрические синапсы — в другие клетки глиального синцития. Такая схема перераспределения внеклеточного калия между областями с разным уровнем нейронной активности косвенно подтверждается в экспериментах с внутриклеточной регистрацией потенциалов глии оптического нерва тритона (Necturus) (рис. 14.16) и Мюллеровских клеток сетчатки саламандры (рис. 14.10).
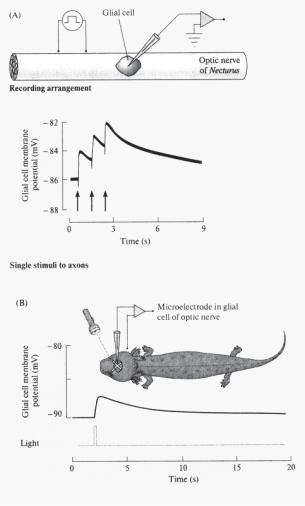
Ðèñ. 14.16. Связь мембранного потенциала глиальной клетки оптического нерва тритона с активностью прилегающих оптических волокон (по: Orkand, Nicholls, Kuffler, 1966). (A) Электрическая стимуляция оптических волокон сопровождается деполяризацией в соседних глиальных клетках. Вверху — схема стимуляции (recording arrangement) и регистрации активности глиальной клетки (glial cell) и оптического нерва тритона (Optic nerve of Necturus), внизу — график зависимости величи- ны мембранного потенциала глиальной клетки (ось ординат, мВ) от частоты электрической стимуляции (моменты нанесения стимулов указаны стрелками). (B) Короткая (0.1 с) вспышка света (light) активирует оптический нерв, что сопровождается деполяризацией в соседней глиальной клетке (glial cell membrane potential, mV).
455
Теоретический анализ функционирования «пространственного буфера», основанный на ряде предположений о пассивном и активном транспорте калия в нейронах и глии, представлен в работе Odette L. L. & Newman E. A. (1988).
Из вышеизложенного следует, что активность глии косвенным образом — через реакции на изменения в распределении внеклеточного калия и поглощение внеклеточного кальция — отражает активность нейронных структур. Согласно широко распространенной среди нейроученых «глиальной гипотезе» Ройтбака А.И., суммарная активность головного мозга (электроэнцефалограмма, ЭЭГ) является отражением электрической активности глии. С этой точки зрения, сигналы глии, а не нейронов, являются до настоящего времени прямым и основным источником информации о работе мозга в нейронауках. Гипотеза Ройтбака А.И. подтверждается в работах на сетчатке («кусочке мозга, вынесенном наружу»), где экспериментально продемонстрирована тесная связь между внутриклеточно регистрируемыми реакциями Мюллеровских клеток и суммарной электрической активностью сетчатки (электроретинограммой, ЭРГ) (Newman, 1985, 1988).
14.3.5. Участие глии в формировании гематоэнцефалического барьера и реакциях иммунной системы
Глия является важной составной частью гематоэнцефалического барьера (ГЭБ) (рис. 14.17).
При этом в ЦНС (головной и спинной мозг) барьерные функции выполняются, в основном, нейроглией (астроциты, олигодендроциты) и сосудистыми сплетениями, а в периферической нервной системе — Шванновскими клетками. В опытах на культуре ткани показано, что плотные контакты между клетками эндотелия кровеносных сосудов, обеспечивающие непроницаемость сосудов для чужеродных молекул, формируются только в присутствии астроцитов (Tao-Cheng et al., 1990). О влиянии глии не только на формирование, но и на функционирование уже сформированного ГЭБ свидетельствуют также данные о регуляции глиальными клетками просвета сосудов путем воздействия на образующие ГЭБ эндотелиальные клетки. По гипотезе Paulson & Newman (1987), такое воздействие осуществляется с помощью ионов калия, протонов или закиси азота (NO), выделяемых из отростков астроцитов.
456

Ðèñ. 14.17. Особенности организации гематоэнцефалического барьера в ЦНС и периферической нервной системе (по: Савельев, 2005). В ЦНС (головной и спинной мозг) барьерные функции выполняются нейроглией (астроциты, олигодендроциты) и сосудистыми сплетениями, а в периферической нервной системе — Шванновскими клетками.
457
Небольшое число работ указывает на возможное участие глии во взаимодействиях между нервной и иммунной системами (Ник. и др., 2003). В культуре ткани и in situ обнаружено, что астроциты могут взаимодействовать с Т-лимфоцитами, а совместные действия микроглии и Т-лимфоцитов могут вызывать острое воспаление мозговой ткани (Newman, Reichenbach, 1996; Newman, Sahs, 1998). Эти данные особенно интересны в свете устоявшихся представлений о независимости нервной системы от контроля со стороны иммунной системы.
14.3.6.Участие глии в развитии
èфункционировании нейронных сетей
14.3.6.1.Глия и нейрогенез.
В процессе эмбрионального развития нервной системы все клетки макроглии секретируют вещества, необходимые для управления ростом нервной ткани, — фактор роста нервов (nerve growth factor, NGF); специфический глиальный белок, стимулирующий рост нейритов (glialderived nexin, GDN): ламинин, белок-репеллент N1-25/250 (Николлс с соавт., 2003). Еще один процесс в развивающейся нервной системе, протекающий с активным участием глии, — это формирование ядерных структур, границы которых еще до момента начала агрегации нейронов в ядра определяются (обозначаются) глиальными клетками (Willbold et al., 1995). И, наконец, миграция развивающихся нейронов к местам назначения осуществляется, как в эмбриогенезе, так и во время прижизненного нейрогенеза, вдоль отростков клеток радиальной глии в ЦНС и отростков Шванновских клеток в периферической нервной системе (рис. 14.11, 14.18) (Rakic, 1981; Hatten, 1999).
То, куда будет мигрировать клетка-предшественник нейрона (в гиппокамп, кору и т.д.), определяется при этом специфичностью контакта клетки с поверхностно расположенными на мембране глиальной клетки молекулами аттрактантов (по принципу «ключ-замок»; Николлс с соавт., 2003). Спраутинг аксонов, стимулируемый глиальными клетками, играет важную роль в процессах восстановления поврежденной нервной ткани во взрослом организме (рис. 14.11–14.13). В опытах на культуре ткани сетчатки крысы выявлено, что синапсы на зрительных нейронах образуются только в присутствии астроцитов, выделяющих при этом такие критически важные соединения, как жировой комплекс под названием àðîÅ/холестерин и белок тромбоспондин (Филдз, 2004).
458
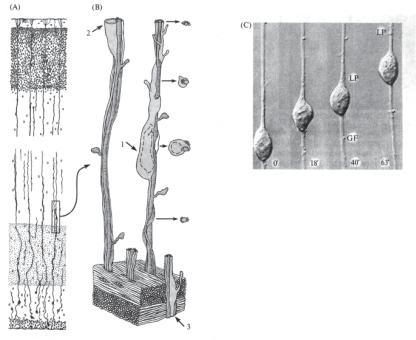
Ðèñ. 14.18. Роль радиальной глии в процессах формирования развивающейся нервной системы (по: Rakic, 1988 (A, B); Hatten, 1990 (C)). (А) На срезе развивающейся затылочной коры плода обезьяны радиальные волокна расположены вдоль путей миграции формирующихся нейронов от вентрикулярной зоны (внизу) к поверхностным слоям (вверху). (B) Клетки коры мигрируют к местам своего назначения с помощью специальных (ведущих) отростков, ориентированных вдоль волокон радиальной глии как своеобразных «направляющих» (проводников). Клетки 1, 2, 3 — вновь образующиеся нейроны на разных этапах миграции из вентрикулярной зоны в поверхностные слои (клетка 3 — в начале, а клетка 2 — в конце своего «путешествия»). (С) Видеосъемка миграции нейрона гиппокампа вдоль волокна радиальной глиальной клетки (glial fiber, GF) in vitro: с течением времени (указано в минутах внизу) ведущий отросток (leading process, LP) постепенно перемещается вверх вместе со следующим за ним телом новой клетки. Однако имеются данные, свидетельствующие о возможности роста и регенерации нервной ткани и вне контакта с глией (Николлс с соавт., 2003). Так, большая часть нейронных синапсов формируется на тех этапах эмбриогенеза, когда количество глиальных клеток крайне мало. Эти и другие факты подобного рода противоречат, по мнению Дж.Николлса с соавт. (2003), устоявшейся точке зрения, согласно которой «синапсы не могут ни формироваться, ни функционировать в отсутствие глиальных клеток» (Pfrieger, Barres, 1996).
459