

В каскадном механизме передачи сигнала заложен механизм усиления сигнала
На рисунке слева показана схема влияния адреналина на гепатоцит. Связывание одной молекулы гормона с одной молекулой рецептора активирует несколько молекул Gsбелков. Каждая активная молекулаGsактивирует молекулу аденилатциклазы, которая катализирует образование многих молекул цАМФ (на рис 40 молекул). цАМФ как вторичный посредник активирует ПKA(10 молекул ПКА, поскольку на активирование каждой молекулы приходится 4 молекулы цАМФ) , каждая молекула которой катализирует фосфорилирование многих (100 молекул ) молекул белка -мишени (киназы фосфорилазыb). Последняя активирует путем фосфорилирования фосфорилазуb(1000 молекул ). И как результат, под влиянием одной молекулы адреналина из клетки печени высвобождается 10000 молекул глюкозы. Несомненно, приведенные данные дают лишь приблизительные представления о количественных изменениях каскадного механизма усиления действия гормона.
Э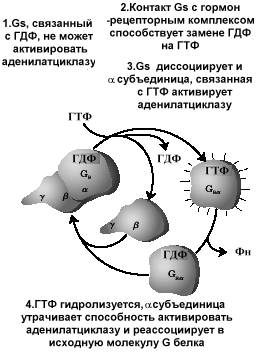 та
киназа в свою очередь активирует
гликогенфофорилазу, которая обеспечивает
быструю мобилизации глюкозы из гликогена.
Конечным результатом влияния такого
последовательного взаимодействия
ферментов является усиление гормонального
сигнала на несколько порядков, что и
объясняет очень низкую концентрацию
адреналина (или любого другого гормона)
необходимую для проявления его активности.
та
киназа в свою очередь активирует
гликогенфофорилазу, которая обеспечивает
быструю мобилизации глюкозы из гликогена.
Конечным результатом влияния такого
последовательного взаимодействия
ферментов является усиление гормонального
сигнала на несколько порядков, что и
объясняет очень низкую концентрацию
адреналина (или любого другого гормона)
необходимую для проявления его активности.
На каждом этапе каскадного механизма усиления сигнала есть свои выключатели.
Столь активное усиление сигнала требует тщательного контроля над количественными параметрами этого усиления. G-белки - один из примеров биологических выключателей, широко используемых в клетке. Связывание гормона и рецептора способствует замене наG–белке ГДФ на ГТФ, что является сигналом для диссоциации субъединицGбелка и активированию субединицами этого белка эффекторных белков ( аденилатциклазы). Продолжительность этого действия определяется скоростью гидролиза ГТФ субъединицей и после превращения ГТФ в ГДФ действие гормон –рецепторного комплекса выключается. Эффект токсина холерного вибриона связан с ингибированием ГТФазной активностиGбелков мембран энтероцитов. При этом естественный выключатель переходит в состояние постоянного включения, что и лежит в основе патологических проявлений холеры.
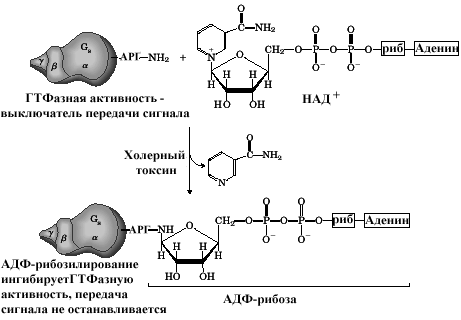
Рис 11.8. Влияние холерного токсина на Gбелок
Активирующее влияние комплекса гормон - рецептор может снижаться и фосфорилированием цитоплазматической части рецептора специфическими протеинкиназами. Как отмечено ранее, системы передачи сигнала подвергаются десенситизации (десенсибилизации) при длительном взаимодействии гормона с рецептором (высокий уровень гормона в межклеточном пространстве). Десенсибилизация -адренэргического рецептора опосредуется протеинкиназой, которая фосфорилирует внутриклеточный домен рецептора, взаимодействующий с Gs.
Эффект одного сигнала может модулирваться влиянием другого сиганала на ту же клетку.Некоторый гормоны действуют путем торможения аденилатциклазы,понижая уровень цАМФи ингибируя фосфорилирование белков. Например, связывание соматостатина со своим рецептором ведет к активированию ингибирующегоG белка илиGi,структурно гомологичногоGs, который ингибирует аденилатциклазу и понижает уровень цАМФ. Соматостатин тем самым уравновешивает эффекты глюкагона.
В жировой ткани, простагландин E1 (ПГЕ1) ингибирует аденилатциклазу, снижая уровень цАМФ и замедляет мобилизацию запасов липидов, вызванных адреналином и глюкагоном. В некоторых других тканях ПГЕ1стимулирует синтез цАМФ, потому что его рецепторы соединены с аденилатциклазой через стимулирующийGsбелок. В тканях с2 -адренэргическим рецептором, адреналин понижает уровень цАМФ, потому что2 рецепторы связаны с аденилатциклазой при участии белкаGi.
Таким образом , внеклеточный сигнал типа адреналина или PGE1 может иметь весьма различные эффекты на различные ткани или клеточные типы, в зависимости от (1) типа рецептора, (2) типа белка G(GsилиGi) с которым рецептор соединен, и (3) набора ферментов мишенейPKAв клетке.
Как упоминалось выше, уровень цАМФ быстро понижается под влиянием фосфодиэстеразы. На уровне ферментов, активность которых регулируется протеинкиназами, функционируют протеинфосфатазы, дефосфорилирующие белки субстраты протеинкиназ и снимающие эффект гормона.