

ЦАмф - первый вторичный посредник в истории исследований переноса сигналов
 Эрл
Уилбер Сазерленд, американский биохимик,
в середине 50-х годов ХХ столетия, при
исследовании механизма активирования
распада гликогена под влиянием адреналина
и глюкагона обнаружил, что эти гормоны
не оказывают непосредственного влияния
на распад гликогена. При инкубации
мембран клеток печени в присутствии
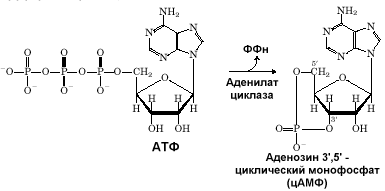
АТФ в среде появлялось термостабильное
соединение, которое Сазерленд
идентифицировал как циклическую
адениловую кислоту (цАМФ). Это соединение
возникало из АТФ под влиянием фермента,
который он назвал аденилатциклазой.
К
Эрл
Уилбер Сазерленд, американский биохимик,
в середине 50-х годов ХХ столетия, при
исследовании механизма активирования
распада гликогена под влиянием адреналина
и глюкагона обнаружил, что эти гормоны
не оказывают непосредственного влияния
на распад гликогена. При инкубации
мембран клеток печени в присутствии
АТФ в среде появлялось термостабильное
соединение, которое Сазерленд
идентифицировал как циклическую
адениловую кислоту (цАМФ). Это соединение
возникало из АТФ под влиянием фермента,
который он назвал аденилатциклазой.
К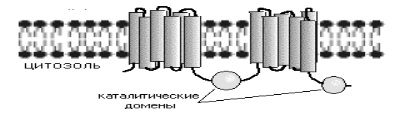 ак
оказалось аденилатциклаза широко
распространенный мембраносвязанный
белок, состоящий из 1100 аминокислот,
которые формируют 2 кластера по 6
трансмембранных сегментов, разделенных
на цитоплазматической стороне двумя
каталитическими доменами. Известны к
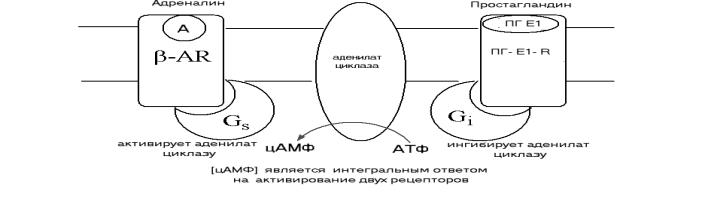
настоящему времени 6 типов Рис.11.6. Влияние
адреналина и простагландина Е1 на
активность аденилатциклазы
ак
оказалось аденилатциклаза широко
распространенный мембраносвязанный
белок, состоящий из 1100 аминокислот,
которые формируют 2 кластера по 6
трансмембранных сегментов, разделенных
на цитоплазматической стороне двумя
каталитическими доменами. Известны к
настоящему времени 6 типов Рис.11.6. Влияние
адреналина и простагландина Е1 на
активность аденилатциклазы
аденилатциклаз. Активность этих ферментов регулируется G- белками. В зависимости от типа аденилатциклаз, они могут взаимодействовать с sсубъединицейGбелка и переходить в активное состояния или с I cубъединицей и ингибироваться.
На рис 11.6 приводится обобщенное представление о интегральном ответе аденилатциклазы. Одна и та же молекула не может взаимодействовать с двумя регуляторами с противоположным действием.
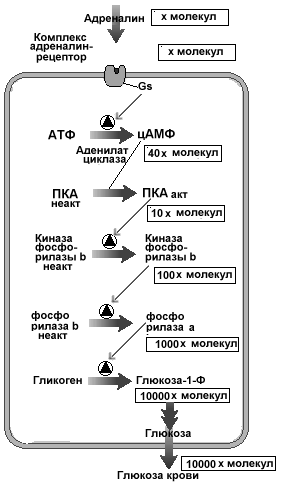
Уровень цАМФ в клетках очень низок (< 10-7М), но при активировании аденилатциклазы может быстро( секунды) увеличиться в несколько раз. Однако это повышение бывает кратковременным. Распад цАМФ катализируется цАМФ-фосфодиэстеразами (ФДЭ). Эти ферменты катализируют гидролиз фосфодиэфирной связи в молекуле цАМФ с образованием аденозин-5-монофосфата. Активность ФДЭ также можно регулировать. Известно, например, что некоторые производные пурина (кофеин) могут быть ингибиторами этого фермента, что также может приводить к повышению уровня цАМФ.
ЦАмф- аллостерический регулятор протеинкиназы а (пка).
 Эффект
усиления распада гликогена, наблюдаемый
Сазерлендом не был результатом прямого
влияния цАМФ на фосфорилазу гликогена.
Активность фосфорилазы изменялась
путем ее фосфорилирования при участии
еще одного фермента ц-АМФ-зависимой
протеинкиназы (протеинкиназы А - ПКА).
ЦАМФ является аллостерическим активатором
протеинкиназы. В последующем было
выделено несколько типов ПКА, имеющих
схожее строение в разных тканях. Это
гетеромеры, состоящие из 2-х каталитических
(С2) и одной регуляторной субъединицы,
состоящей из двух полипептидных цепей
(R2).
Эффект
усиления распада гликогена, наблюдаемый
Сазерлендом не был результатом прямого
влияния цАМФ на фосфорилазу гликогена.
Активность фосфорилазы изменялась
путем ее фосфорилирования при участии
еще одного фермента ц-АМФ-зависимой
протеинкиназы (протеинкиназы А - ПКА).
ЦАМФ является аллостерическим активатором
протеинкиназы. В последующем было
выделено несколько типов ПКА, имеющих
схожее строение в разных тканях. Это
гетеромеры, состоящие из 2-х каталитических
(С2) и одной регуляторной субъединицы,
состоящей из двух полипептидных цепей
(R2).
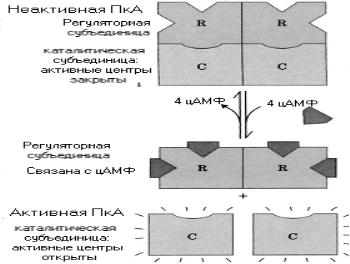
Были выделены 3 типа каталитических () субъединиц и два типа регуляторных (I и II). Холоэнзимы R2С2неактивны. Присоединение цАМФ вызывает диссоциацию фермента. При этом каталитические субъединицы освобождаются и начинают активно катализировать перенос фосфорного остатка с АТФ на определенные остатки серина или треонина некоторых белков органа- мишени. Обычно специфичекой последовательностью аминокислот участков фосфорилирования АРГ-АРГ-Х-СЕР или ЛИЗ-АРГ-Х-Х-СЕР, где Х- любая аминокислота.
Такая ковалентная модификация фермента стабилизирует одно из конформационных состояний фермента: активное -расслабленное или неактивное- напряженное. Это уже зависит от свойств данного фермента. Так, фосфорилирование гликогенфосфорилазы активирует ее, а гликогенсинтетазы, наоборот, тормозит ее активность. Эффекты цАМФ обычно непродолжительны. В клетках имеются ферменты дефосфорилирующие белки, фосфорилируемые протеинкиназой А. Известны две главные протеинфосфатазы., одна из которых сама регулируется цАМФ. Последняя наиболее активна в отсутствии цАМФ.
Р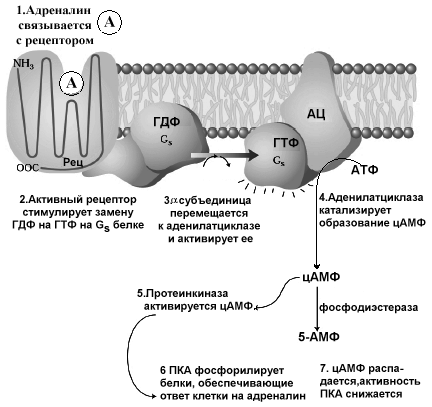 ис11.7.
Схема этапов влияния адреналина на
клетку
ис11.7.
Схема этапов влияния адреналина на
клетку
цАМФ может выполнять регуляторные функции и другими альтернативными механизмами, например, открытием цАМФ-зависимых ионных каналов. На рис 11.7 суммированы данные о механизме действия адреналина на адренэргический рецептор
|
Таблица 11.4.Некоторые клеточные ответы на действие гормонов, опосредуемое циклическим АМФ | ||
|
Гормон |
Клетки-мишени |
Главный ответ |
|
Тиротропин |
Тироциты |
Синтез и секреция тироксина |
|
Кортикотропин |
Клетки пучковой и сетчатой зоны коры надпочечников |
Секреция кортизола |
|
Лютропин |
Клетки Лейдига яичников |
Секреция прогестерона |
|
Адреналин |
Миоциты, гепатоциты |
Распад гликогена |
|
Паратгормон |
Остеокласты |
Резорбция кости |
|
Адреналин |
Р-клетки водителя ритма сердца |
Увеличение частоты сокращений |
|
Вазопрессин |
Эпителиоциты канальцев нефрона почки |
Реабсорбция воды |
|
Адреналин, кортикотропин, глюкагон, тиротропин |
Адипоциты |
Расщепление триацилглицеролов |
В таблице 11.4 представлены некоторые клеточные ответы на действие гормонов, активирующих аденилатциклазный путь сигнализации. Важно отметить, что разные лиганды, действующие через различные рецепторы на аденилатциклазу в данной клетке, обычно вызывают одну и ту же ответную реакцию. Например, в адипоцитах аденилатциклазу активируют по крайней мере четыре разных гормона, (адреналин, кортикотропин, глюкагон, тиротропин) и все они вызывают расщепление триацилглицеролов (резервная форма жира) до жирных кислот. Похоже, что различные рецепторы для этих гормонов активируют общий пул молекул аденилатциклазы.
 В
некоторых животных клетках повышение
уровня цАМФ приводит к активации
транскрипции определенных генов.
Например, в нейроэндокринных клетках
гипоталамуса цАМФ "включает" ген,
кодирующий синтез пептидного гормона
соматостатина. Каков же механизм этого
процесса? Дело в том, что активированная
каталитическая субъединица ПКА может
траслоцироваться из цитоплазмы в ядро.
На уровне ядерного компартмента влияние
протеинкиназы А реализуется через
фосфорилирование так называемых
ген-регуляторных белков (generegulatoryproteins),
которые, активируясь, связываются со
специфическими отвечающими элементами
(responseelements),
представляющими собой короткие участки
ДНК строго определенной последовательности
генов-регуляторов, формируя при этом
пусковое звено активации или репрессии
транскрипции соответствующего
транскриптона.
В
некоторых животных клетках повышение
уровня цАМФ приводит к активации
транскрипции определенных генов.
Например, в нейроэндокринных клетках
гипоталамуса цАМФ "включает" ген,
кодирующий синтез пептидного гормона
соматостатина. Каков же механизм этого
процесса? Дело в том, что активированная
каталитическая субъединица ПКА может
траслоцироваться из цитоплазмы в ядро.
На уровне ядерного компартмента влияние
протеинкиназы А реализуется через
фосфорилирование так называемых
ген-регуляторных белков (generegulatoryproteins),
которые, активируясь, связываются со
специфическими отвечающими элементами
(responseelements),
представляющими собой короткие участки
ДНК строго определенной последовательности
генов-регуляторов, формируя при этом
пусковое звено активации или репрессии
транскрипции соответствующего
транскриптона.