

Наиболее широко распространенная группа 1тмс рецепторов в клетках – рецепторные тирозинкиназы (ртк).
В 1982 году впервые был открыт рецептор со свойствами тирозин -специфической протеинкиназы для эпидермального фактора роста (ЭФР). ЭФР- это маленький белок (53 аминокислоты), стимулирующий пролиферацию эпидермальных и некоторых других клеток. Его рецептор - классический 1TMS –рецептор. Он состоит из 1200 аминокислот и большой гликозилированной внеклеточной части, которая обеспечивает связывание ЭФР. При присоединении ЭФР активируется внутриклеточный домен, обладающий свойствами тирозинкиназы. Этот домен рецептора катализирует перенос остатка фосфорной кислоты на тирозиновый остаток самого рецептора и/или на тирозиновый остаток специфического внутриклеточного белка. Как оказалось, рецепторы многих других факторов роста также обладают свойствами тирозинкиназы и имеют ряд общих структурных и функциональных свойств. Можно выделить 4 основные домена в их структуре
Гликозилированный внеклеточный домен, связывающий лиганд.
Внутриклеточный домен с тирозинкиназной активностью.
Внутриклеточный регулирующий домен.
Трансмембранный домен
Аминокислотные последовательности тирозинкиназных доменов РТК высоко консервативны и подобны цАМФ зависимой протеинкиназе (ПкА)в областях связывания АТФ и субстратов. Некоторые РТК имеет вставки аминокислотных последовательностей в киназные домены названные киназными вставками. РТК классифицируются на семейства, на основе структурных особенностей их внеклеточных частей: наличие или отсутствие киназных вставок, богатых цистеинами последовательностей, иммуноглобулинподобных областей, лейцин – богатых областей и т.д. Выделяют на по крайней мере на 14 различных семейств РПК.
Некоторые рецепторные протеинкиназы могут катализировать фосфорилирование специальных белков цитозоля. Примером такого рода рецепторов является рецептор инсулина. Присоединение инсулина вызывает активирование его киназного домена, который помимо аутофосфорилирования, катализирует фосфорилирование остатков тирозина специального белка , названного субстратом инсулинового рецептора IRS-1.
Фосфорилированные тирозиновые остатки связываются со специфическими доменами белков.
Фосфотирозины , образующиеся в белках под влиянием рецеторной протеинкиназы затем служат местом присоединения и активирования многочисленных внутриклеточных белков.
Способность связываться с фосфорилированными тирозинами обусловлена специфическими доменами в структуре таких белков, названными SH2 иSH3 доменами. Впервые эти домены бы
ли обнаружены при исследовании механизмов развития опухолей (саркомы). Семейство протоонкогенов, выделенное при этих исследованиях и обладающих способностью связываться с тирозинами, получило название Src-семейства. Гомологичные домены, обнаруженные в других белках получили названиеSH2 и SH3 по аналогии с соответствующими участкамиSrcбелков ((S)rc(H)omologydomain2 или 3 ).
Среди таких белков можно назвать фосфатидилинозитол 3-киназу (ФИ-3), фосфолипазу С-белок, активирующий ГТФазу (GAP), Src –подобную нерецепторную тирозинкиназу и многие другие.
Существует группа белков, названных малыми SHадапторными белками и состоящими из 2 доменовSH2 иSH3 . Эти белки не обладают каталитической активностью, а выполняют связующую роль между активированными тирозинкиназами и другими белками, которые не имеют своихSH2 и SH3доменов.Известны и другие структуры, обеспечивающие узнавание фосфорилированных молекул.
Фосфорилированные тирозины, таким образом, являясь местом присоединения разнообразных белков, служат инициаторами целых каскадов реакций, направленных на усиление сигналов и специфическое включение или выключение определенных функций, необходимых для выживания клеток, их пролиферации или программированной гибели.
В передаче сигнала с участием 1ТМС рецепторов тоже есть выключатели.
В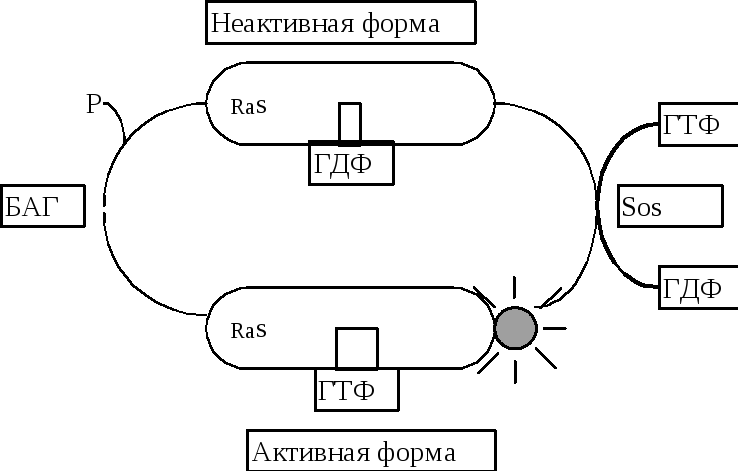 ажную
роль в в передаче сигналов с участием
активных рецепторных тирозинкиназ
играют Ras белки. Ras белки - это семейство
мономерных ГТФаз. ПодобноG-белкам
(тримерным ГТФазам), они содержат на
С-конце ковалентно связанный фарнезильный
или геранильный остаток. С помощью
такого гидрофобного конца белки Ras
(p21ras) прикрепляются к к цитоплазматической
стороне плазматической мембраны.
ажную
роль в в передаче сигналов с участием
активных рецепторных тирозинкиназ
играют Ras белки. Ras белки - это семейство
мономерных ГТФаз. ПодобноG-белкам
(тримерным ГТФазам), они содержат на
С-конце ковалентно связанный фарнезильный
или геранильный остаток. С помощью
такого гидрофобного конца белки Ras
(p21ras) прикрепляются к к цитоплазматической
стороне плазматической мембраны.
Ras белки участвуют в передаче сигнала от рецеторной тирозинкиназы к ядру при стимуляции клеточной пролиферации или дифференцировки факторами роста или гормонами. Если эти белки связать антителами, то нарушается передача информации и, как следствие, нарушается дифференцировка клеток , которая обычно вызывается активированием рецепторных тирозинкиназ. Как подобает мономерным ГТФазам, Ras белки могут переключать свою конформацию и быть активными, если к ним присоединяется ГТФ и становиться неактивными при замене ГТФ на ГДФ. Правда, Ras белки катализируют гидролиз ГТФ в 100 раз медленнее, чем большие тримерные G- белки и, учитывая высокую концентрацию ГТФ в клетке, Ras белки могут находиться в активном состоянии долго. Способность Ras белков взаимодействовать с ГТФ или катализировать ее гидролиз в свою очередь регулируется дополнительными белками. В клетках существуют 2 класса белков такого рода. Белок, активирующий ГТФазу (БАГ) усиливает степень гидролиза ГТФ Ras белками и поэтому инактивирует их, ингибируя передачу сигнала .
Этому негативному регулятору противостоит белок, который ускоряет освобождение связанного ГДФ и замену его на ГТФ из цитозоля и поэтому оказывает активирующий эффект на Ras белок. Этот белок называют фактором, высвобождающим гуаниловые нуклеотиды (guanylnucleotide relisyng protein GRP. На рис 12-15 он обозначен Sos). Ras белки и их регуляторы консервативны в эволюции и подобны по строению у разных видов живых организмов.
Фосфорилирование тирозина, катализируемое рецепторной протеинкиназой на цитоплазматической поверхности мембраны и активирование Ras длится короткий промежуток времени: специфические протеинфосфатазы катализируют дефосфорилирование тирозина, а активный Ras инактивируется путем гидролиза ГТФ, приостанавливая передачу сигнала внутрь клетки.