

Пассивный транспорт воды
Не существует молекулярных механизмов активного транспорта воды, подобных мембранному белку, активно транспортирующему Na+, например Na+,К+-АТФаза. Вода может перемещаться либо по градиенту гидростатического, либо по градиенту осмотического давления, и поскольку гидростатический градиент через эпителий кишечника равен нулю, то основным механизмом транспорта воды является осмотический градиент.
Термин "осмотическое давление" был долгое время не очень понятным для студентов и практических врачей как минимум по двум причинам. Во-первых, он означает, что существует еще какое-то давление, помимо гидростатического. На самом деле "осмотическое давление" является неправильным термином, который появился только из-за того, что градиент растворенных веществ повышает гидростатическое давление в осмометре. Лучшим названием было бы "осмотический потенциал".
Осмотическое давление , обычно выражается как
= RTCs,
где: Cs — общая концентрация растворенных веществ в осмолях/кг Н2О. Разница в осмотических давлениях через кишечный эпителий выражается как , то есть
= RTCs.
Здесь мы видим вторую причину неясности термина "осмотическое давление". является пассивной силой, возникающей из-за разницы в концентрации воды, но для удобства выражается в виде разницы в концентрации солей. Поэтому необходимо понимать, что градиент растворенных веществ определяет градиент концентрации воды; оба этих градиента определяют реальную направленность движения воды из зоны ее высокой концентрации в зоны с высокой концентрацией растворенных веществ.
Еще одно важное обстоятельство заключается в том, что общий осмотический эффект растворенных веществ более выражен, если это растворенное вещество находится в каком-то ограниченном объеме. Например, при непереносимости лактозы неабсорбированная глюкоза не может покинуть просвет кишечника и создает выраженный осмотический эффект, вызывая осмотическую диарею. Понятие осмотической диареи основано на том факте, что причиной секреции воды в некоторых случаях является неабсорбируемый осмотический эквивалент. Необходимо также помнить, что все перемещения воды происходят посредством осмотических механизмов, хотя причины осмотических нарушений (как и при секреторной диарее) не всегда понятны.
Слизистая оболочка тонкой кишки легко проницаема для воды
Перемещение больших объемов воды через стенку кишечника предполагает, что эпителий кишечника легко для нее проницаем, поскольку через малопроницаемый барьер даже при высоком осмотическом градиенте перемещения молекул воды не происходит. Принципы, по которым действуют механизмы транспорта воды через мембраны, аналогичны закону Ома для электричества: поток воды пропорционален осмотическому давлению и степени проницаемости мембраны для воды.
Подобно другим веществам, проникающим через кишечный эпителий, вода проходит либо трансцеллюлярно, либо через межклеточные соединения и пространства. Молекулярные механизмы этих путей транспорта воды недостаточно хорошо изучены, однако знание действия подобных механизмов в других органах, например в почках, позволяет проводить определенные аналогии. Исследование простого липидного бислоя, являющегося нормальным структурным компонентом мембраны многих клеток, показало, что он достаточно хорошо проницаем для воды. Поэтому некоторое время считали, что транспорт воды происходит через липидный слой клетки. Однако в последнее время получены данные о существовании в клеточной мембране особого класса белков, служащих каналами для проведения воды. Возможно эти белки, или что-то похожее на них, могут отвечать за проницаемость энтероцитов для воды. Еще меньше изучены механизмы межклеточного транспорта воды, хотя известно, что часть воды проходит именно этим путем (рис. 5-5).

Рис. 5-5. Трансцеллюлярный и интерцеллюлярный пути транспорта воды
Транспорт солей определяет транспорт воды
Еще в ранних работах по исследованию транспорта веществ в кишечнике показано, что в норме абсорбция воды и электролитов происходит таким образом, что, несмотря на большие объемы перемещения электролитов и воды, содержимое кишечника остается изотоничным плазме крови. Эти данные не позволили прежним исследователям объяснить, как происходит всасывание воды, если нет градиента осмотического давления, поэтому казалось, что существует механизм активного транспорта воды в кишечнике. Позже было показано, что всасывание воды происходит исключительно пассивно посредством осмотического градиента, возникающего вследствие всасывания электролитов, и что оба этих процесса тесно связаны. Отсутствие реальной разницы осмотических давлений в стенке кишечника является следствием действия двух факторов. Первый — высокая проницаемость стенки кишечника для воды предполагает настолько маленький трансэпителиальный осмотический градиент (), необходимый для всасывания воды, что его трудно определить экспериментально. Второй, по-видимому, связан с тем, что в межклеточных латеральных пространствах происходит кумуляция солей, что создает местный локальный градиент, способствующий перемещению жидкости. Главный принцип остается прежним — всасывание воды происходит пассивно вследствие транспорта солей, преимущественно NaCl. Это является ключом к пониманию того, что именно нарушение транспорта солей — основная причина многих видов диареи.
Секреция жидкости в кишечнике также регулируется секрецией солей клетками, расположенными в криптах. Термин "секреторная диарея" является следствием того факта, что бактериальные токсины и другие агенты могут вызывать повышенную стимуляцию секреции солей в просвет кишечника с соответствующим объемом секреции воды. Причина этого — нарушение молекулярных механизмов контроля, которые в норме регулируют кишечную секрецию.
Поверхностные клетки и клетки крипт специализированы для абсорбции и секреции
Абсорбция и секреция солей, приводящие к перемещению воды через стенку тонкой кишки, осуществляются специализированными клетками эпителия. Абсорбция осуществляется эпителиальными клетками ворсинок, которые активно абсорбируют Na+. Секреция производится клетками, расположенными в криптах, которые активно перемещают Cl–. Несмотря на то, что эти клетки активно транспортируют столь разные ионы, параллельно осуществляется и котранспорт: абсорбция Na+ не может происходить без абсорбции Cl–, а секреция Cl– происходит с параллельным перемещением Na+. Активный транспорт является основой перемещения ионов, а перемещение ионов с противоположным зарядом — также неотъемлемая часть этого процесса.
На Рис. 5-6 показаны молекулярные механизмы транспорта солей клетками ворсинок и крипт в тонкой кишке. В обоих типах клеток вектор транспорта ионов связан с выраженным полярным распределением мембранных белков, которые способствуют транспорту ионов через апикальные и базолатеральные участки мембраны клеток. В настоящее время найдены и специальные гены, ответственные за синтез этих белков.
Особой структурой как в абсорбирующих, так и в секретирующих клетках является Na+,K+-ATФaзa. Этот транспортный белок расположен исключительно на базолатеральных участках мембраны клеток кишечника и связывает непосредственно процессы ионного транспорта и энергетические процессы в клетках. Na+,K+-АТФаза поддерживает низкий уровень внутриклеточного Na+ и высокую концентрацию внутриклеточного К+ за счет гидролиза АТФ. Na+,K+-ATФaзa создает градиенты концентраций Na+ и K+, необходимые для поддержания абсорбционной и секреторной активности в эпителиальных клетках кишечника.
Абсорбирующие клетки
Мембрана щеточной каемки абсорбирующих клеток специализирована в отношении пассивного транспорта Na+ в клетки из просвета кишки. В тонкой кишке это происходит преимущественно двумя путями. Первый путь: антипорт Na+/H+, специализирующийся на катализе обмена этих ионов в соотношении 1:1. При этом ионы натрия входят в клетку, а протоны водорода выходят из клетки. Второй механизм — котранспортер Na+/глюкоза, переносящий внутрь клетки ионы Na+ и молекулы глюкозы в соотношении 1:1, либо в соотношении 2:1. Таким образом, в присутствии глюкозы Na+ поступает в клетку вместе с ней. В этом механизме свободная энергия натриевого градиента способствует абсорбции глюкозы. Однако необходимо отметить, что наличие глюкозы в просвете кишки резко повышает абсорб
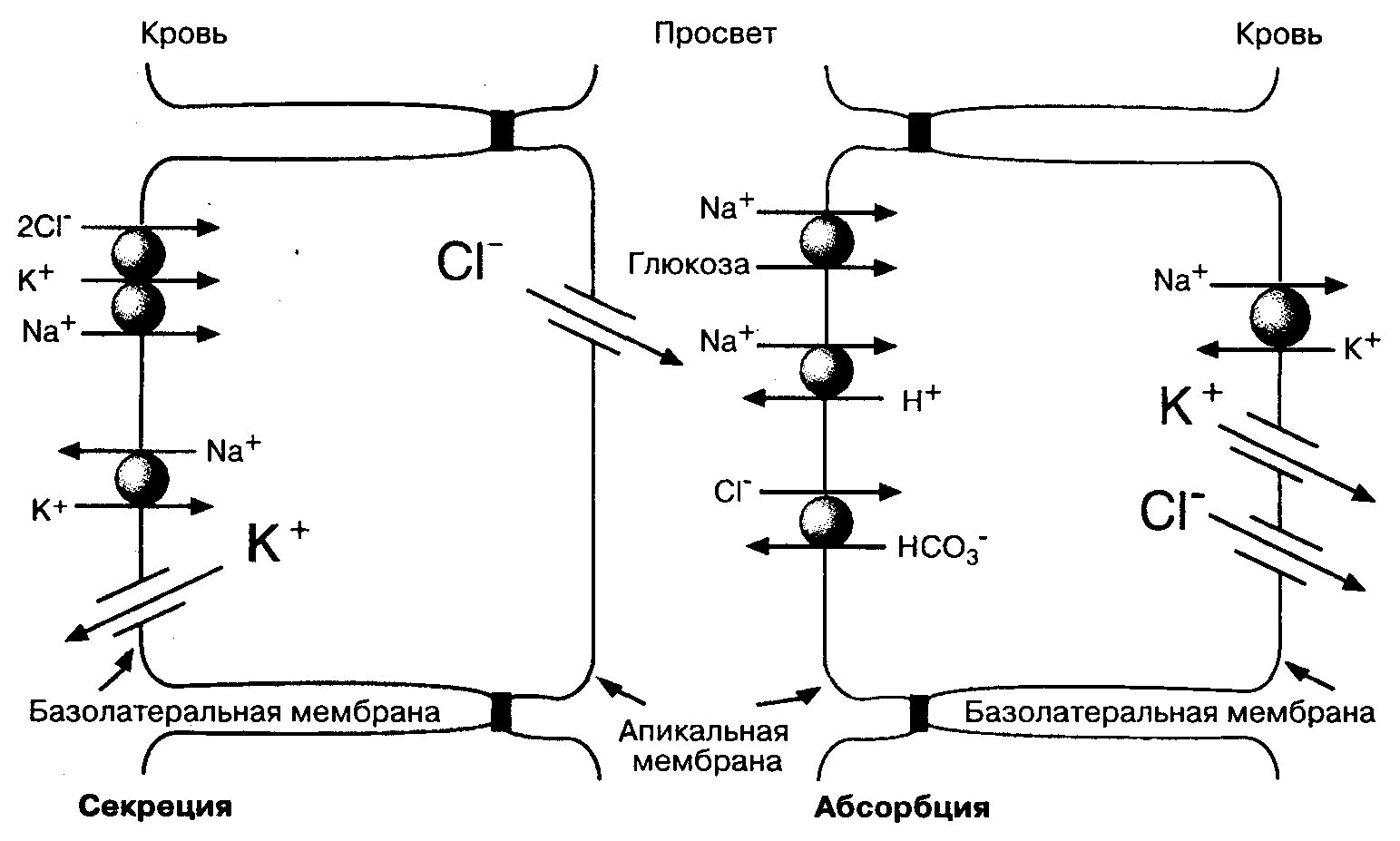
Рис. 5-6. Молекулярные механизмы транспорта веществ клетками ворсинок и крипт в тонкой кишке. Показаны некоторые мембранные белки, участвующие в транспорте солей через апикальные и базолатеральные участки мембраны. Разница электрических потенциалов (Vm) между апикальными и базолатеральными участками мембран абсорбирующих и секретирующих клеток ориентирована таким образом, что клетки относительно экстрацеллюлярной жидкости имеют отрицательный заряд. Величина Vm на базолатеральном участке мембраны больше, чем Vm апикального участка мембраны, поэтому трансэпителиальный электропотснциал, измеренный через слой эпителиальных клеток, является отрицательным со стороны просвета кишки
цию натрия, усиливая его проникновение в клетки. Этот механизм лежит в основе использования растворов глюкозы перорально для регидратации больных с диареей, так как глюкоза повышает абсорбцию Na+ и воды. Растворы, содержащие только необходимые электролиты, менее эффективны, поскольку они не стимулируют глюкозозависимый транспорт Na+ в клетки.
Na+,K+-ATФaзa базолатерального участка мембраны и белки-переносчики Na+ в апикальных отделах мембраны клеток отвечают за трансцеллюлярный перенос Na+ из просвета кишки в кровь; поэтому активный транспорт Na+ — основной механизм абсорбции солей и воды. Абсорбция Сl–, соответствующая абсорбции натрия, происходит двумя путями. Часть Сl– абсорбируется из межклеточного пространства благодаря слабому электрическому потенциалу, отрицательному со стороны просвета кишки. Кроме этого, существует и трансцеллюлярный механизм абсорбции Сl–, обеспечиваемый Сl–/НСО3–-антипортом в апикальном участке мембраны. Механизм транспорта хлора через базолатеральный участок мембраны пока не совсем понятен: это может быть либо Сl–-селективный канал, либо Сl–,K+-котранспортер. Образование Н+ и НСО3– из угольной кислоты с помощью карбоангидразы способствует сочетанному транспорту Na+ и Сl–.