

2 .
ТЕХНОЛОГИЧЕСКИЙ РАЗДЕЛ
.
ТЕХНОЛОГИЧЕСКИЙ РАЗДЕЛ
2.1. Аналитический обзор и патентная проработка
2.1.1. Механизм реакции гидролиза полисахаридов.
Сущность реакции гидролиза полисахаридов заключается во взаимодействии этих веществ с водой, в результате которого происходит разрыв ацетальных связей между звеньями макромолекул с присоединением по месту разрыва водородного и гидроксильного ионов воды. В результате такого процесса снижается степень полимеризации полисахаридов и в качестве конечного продукта образуются простые сахара.
В общей форме реакция гидролиза целлюлозы может быть выражена следующим уравнением:
(С6Н1005)n +(n-1)Н20 = nС6Н1206.
В производственных условиях гидролиз осуществляется в кислой среде, причем водородные ионы служат катализаторами этой реакции. Ранее считалось, что водородные ионы, находясь в воде в виде ионов гидроксония (Н+3О), могут образовывать с молекулами полисахарида активный нестойкий комплекс. Это соединение деформирует электронное облако ацетальной связи и этим уменьшает ее стойкость. Если возбуждение достигает величины, превышающей силу данной связи, последняя разрывается с присоединением водородного и гидроксильного ионов воды. Освобождающийся при этом ион водорода моментально реагирует с молекулой воды и вновь образует ион гидроксония, который продолжает участвовать в дальнейшей реакции. Путем исследования механизма кислотного гидролиза при помощи метода меченых атомов с применением О18 показана возможность присоединения к гликозидному атому кислорода непосредственно протона Н+ с образованием сопряженной кислоты. Последующий разрыв этой связи происходит с присоединением воды и отщеплением протона по следующей схеме.
Под действием иона гидроксония ацетальный кислород быстро протонизируется (I), ослабленная ацетальная связь рвется с образованием иона карбония (II), который реагирует с водой, давая конечную группу и протон (III). Освободившийся протон с водой вновь образует ион гидроксония (IV).
Наиболее простая реакция гидролиза происходит при превращении дисахаридов (сахарозы, мальтозы, целлобиозы) в простые сахара. В этом случае один акт разрыва связи ведет к образованию конечного продукта. Такие реакции легко протекают при кипячении водных растворов этих веществ в присутствии небольшого количества кислоты. Большинство природных полисахаридов, таких, как целлюлоза, крахмал и ксилан, не растворимы в холодной воде. Однако частично гидролизованные крах-мал и ксилан хорошо растворяются в воде, и в этом случае гидролиз их может быть легко осуществлен в гомогенных условиях, подобно гидролизу дисахаридов.
Механизм реакции гидролиза

Рис 2.1
Весьма трудно путем прямого воздействия разбавленных кислот перевести в растворенное состояние целлюлозу. Однако в некоторых случаях этого удается достичь косвенным путем. Так, раствор целлюлозы можно получить путем предварительного растворения ее в концентрированной кислоте и затем небольшого разбавления полученного раствора водой (при большом разбавлении такая целлюлоза выпадает в осадок.)
При гидролизе полисахаридов в гомогенной среде ацетальные связи в цепи макромолекул разрываются постепенно, причем в любом месте, в результате чего происходит непрерывное уменьшение степени полимеризации полисахаридов. О скорости такой реакции судят или по изменению степени полимеризации полисахарида, или по количеству редуцирующих веществ, перешедших в раствор.
Реакция гидролиза полисахаридов в гетерогенных условиях имеет более сложный механизм, так как здесь решающую роль начинает играть надмолекулярная структура полисахарида и величина межмолекулярных сил. Главной характерной особенностью этого процесса является резкое понижение скорости реакции. Например, скорость гидролиза целлюлозы, протекающего в гетерогенных условиях при температуре 80° и выше, в 300 - 400 раз меньше скорости гидролиза любого полисахарида, находящегося в растворенном состоянии/2/.
2.1.2. Механизм брожения.
Известны многие виды брожения: спиртовое брожение, молочнокислое, пропионовокислое, ацетоно-бутиловое и другие виды. Так как процесс гидролиза связан со спиртовым брожением, то мы его и рассмотрим.
Основные возбудители спиртового брожения – дрожжи играют большую роль в жизни человека. Дрожжи традиционно используют в хлебопечении, для получения спирта и многих других продуктов. По значимости для народного хозяйства с ними могут конкурировать только молочнокислые бактерии. Пожалуй, нет на земном шаре ни одного человека, который бы в своей повседневной жизни не пользовался трудами этих микроорганизмов.
На Востоке в качестве возбудителя спиртового брожения при производстве рисового пива (сакэ) применяется Asperglllus oryzae. Спиртовое брожение могут вызывать также некоторые бактерии (Zymomonas mobilis, Z. anaerobica, Sarcina ventricula, Erwinia amylovora). Однако получение спирта с помощью этил микроорганизмов существенного промышленного значения пока не имеет.
Термином «дрожжи» обозначают одноклеточные эукариотные микроорганизмы, которые в зависимости от наличия и типа полового процесса относят к трем классам грибов: Ascomycetei, Basidiomycetes и Deuteromycetes. Термин «дрожжи» в строгом смысле не имеет таксономического значения.
К классу Ascomycetes относят дрожжи, образующие при половом размножении сумки (аски) с эндогенными спорами, К нему принадлежат представители родов дрожжей, используемых в бродильных производствах, Saccharomyces и Shizosacc haromyces. Класс Basidiomycetes включает дрожжи, формирующие телиоспоры (телейтоспоры) и базидиоподобные спорофоры с экзогенными половыми спорами (споридиями). К Deuteromycetes, или несовершенным грибам, относят дрожжи, у которых не обнаружен половой цикл. Считают, что организмы, принадлежащие к этой группе, произошли от высших грибов в результате утраты ими половых функций.
В процессе эволюции дрожжи хорошо приспособились к обитанию в различных местах, содержащих чаще всего углеводы, Они растут на поверхности сладких плодов, в нектаре цветком, в сокотечениях деревьев, на поверхности листьев, в лесной подстилке и почве. Встречаются дрожжи и в водоемах. Содержатся они в пищеварительном тракте человека и животных. Большинство дрожжей сапрофиты, но среди видов, находящими во внутренних органах и на кожных покровах человека, имеются патогенные или условно патогенные формы, например возбудитель кандидомикозов- Candida albicans. Некоторые дрожжи вызывают болезни растений.
Схема анаэробного разложения углеводов (фруктозо-бис-фосфатный путь)
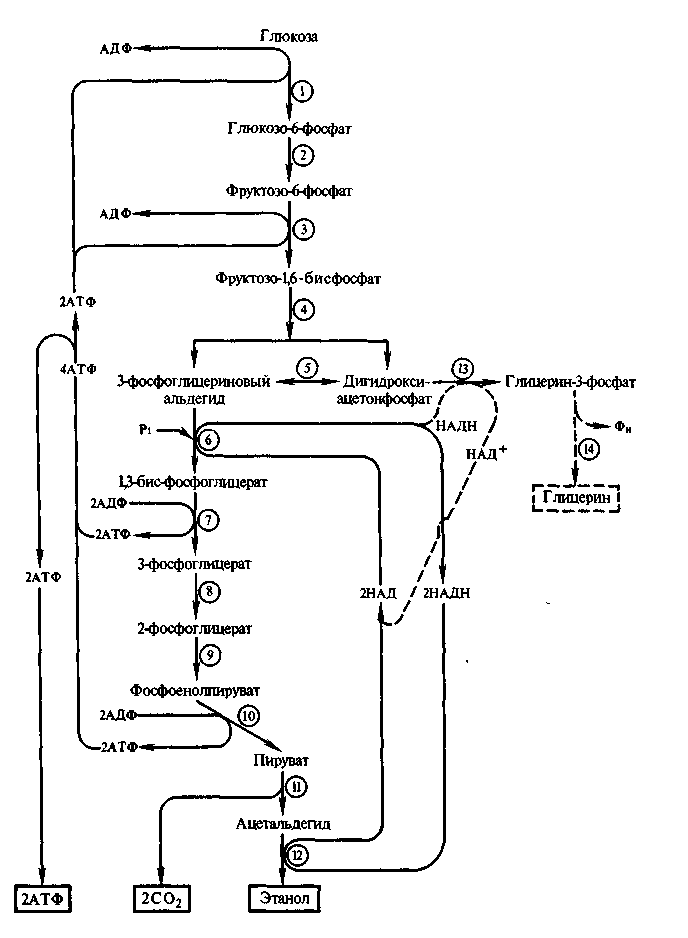
1 - гексокиназа, 2 - глюкозофосфатизомераза, 3 - фосфофруктокиназа, 4- альдолаза, 5 - триозофосфат-изомераза, 6 - глицеральдегидтрифосфатизомираэа,7- фосфоглицераткиназа, 8 - фосфогли-церомутаза, 9 - енолаза, 10 -пируваткиназа, 11 - пируватдекарбоксилаза, 12 - алкогольдегидрогеназа, 13 - глицерофосфатдегидрогеназа, 14 - фосфатаза
Рис. 2.2
Из соединений углерода дрожжи, как правило, лучше используют гексозы. Некоторые виды хорошо растут на средах с пентозами. Из полисахаридов чаще всего утилизируют инулин и крахмал. Известны дрожжи, растущие на средах с углеводородами и некоторыми спиртами, в том числе метанолом и этанолом, а также органическими кислотами и другими углеродными субстратами.
В качестве источника азота дрожжи используют обычно соли аммония, аминокислоты, небольшие пептиды, реже нитраты и нитриты. Некоторые виды нуждаются в одном или более вита минах (чаще в биотине и тиамине), другие -способны все необходимые для роста витамины синтезировать сами.
Большинство дрожжей растет в границах рН от 3,0 до 8,0, оптимальные значения рН от 3,5 до 6,5. Общий диапазон температур для роста дрожжей довольно широк: от 0 (даже -7 0С) до 48-50 °С. Оптимальная температура для роста большинсто видов 28-30 °С, но некоторые расы дрожжей, например, используемые в пивоварении, имеют более низкий температурный оптимум. Известны также облигатно психрофильные дрожжи, не растущие при температуре выше 18-20 °С. Многие дрожжи факультативные анаэробы. В условиях анаэробиоза они получи ют энергию в результате сбраживания углеводов, а в присутствии молекулярного кислорода - за счет аэробного дыхания /5/.
Спиртовое брожение у дрожжей до образования пировиноградной кислоты отличается от гликолиза у высших организме лишь последними этапами, на которых вместо молочной кислоты образуется этиловый спирт. Обусловлено это наличием у дрожжей пируватдекарбоксилазы, катализирующей превращение пирувата в ацетальдегид, который затем восстанавливается в этанол.
Гликолитическим путем (или путем Эмбдена - Мейергофа - Парнаса) его также называют фруктозобисфосфатным (ФБФ-путь), осуществляется разложение глюкозы, галактозы, фруктозы и маннозы. Олигосахариды вначале гидролизуются соответствующими ферментами до гексоз.
По-разному способны утилизировать дрожжи трисахарид раффинозу.
Существовало мнение, что дрожжи используют пентозы лишь в анаэробных условиях. В последнее время установлено, что некоторые из них способны к росту в анаэробных условиях на средах, содержащих ксилозу или ксилулозу; последние подвергаются брожению с образованием этанола. Это имеет важное практическое значение для производств, перерабатывающих в спирт древесину и отходы сельскохозяйственных растений.
Разложение пентоз и высших спиртов осуществляется дрожжами через пентозофосфатный и ФБФ-пути. Спирты вначале дегидрируются до соответствующих гексоз и пентоз.
Некоторые дрожжи (Rhodotorula, Sporobolomyces, Cryptocoteus) используют сахара только в аэробных условиях. Неспособность сбраживать глюкозу связывают с отсутствием у этих организмов пируватдекарбоксилазы или НАД-зависимой алкоголь-дегидрогеназы — ферментов, осуществляющих превращение пирувата в этанол.
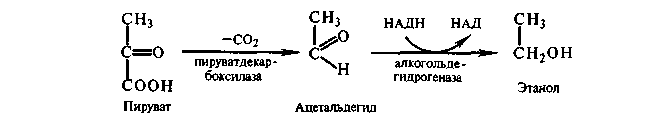
Брожение предполагает строгое равновесие процессов окисления и восстановления. Поэтому НАД, восстановленный на одном на этапов брожения, должен окисляться на другом этапе. Окисление НАД происходит одновременно с восстановлением ацетальдегида в этанол. Такой процесс Нейберг назвал первой формой брожения. Суммарная реакция его:
Глюкоза 2СО2 + 2 этанол
В начале спиртового брожения преобладает глицеринопировиноградное брожение, приводящее к образованию глицерина и пировиноградной кислоты. Однако пировиноградная кислота обнаруживается, как правило, в небольших количествах, поскольку основная ее часть идет на образование различных вторичных продуктов. К ним относятся уксусная, молочная, янтарная, пропионовая, муравьиная и некоторые другие кислоты, ацетон, диацетил альдегиды и сложные эфиры /1/.
Кроме вторичных продуктов при спиртовом брожении образуются побочные продукты - высшие спирты, известные под названием сивушных масел. Почти половину общего количества высших спиртов составляют два изоамиловых спирта: 3-метилбутанол-(1) и 2-метилбутанол-(1). Эти продукты синтезируются из соответствующих кетокислот, образующихся в результате метаболизма углеводов, или из аминокислот. Поэтому вторичные и побочные продукты нельзя строго разграничить. Они существенно влияют на вкус и аромат готового продукта. Образование высших спиртов включает дезаминирование линокислоты в кетокислоту, которая декарбоксилируется в альдегид; последний восстанавливается в спирт. Однако не все высшие спирты синтезируются из аминокислот. Второй путь их образования можно рассматривать как биосинтез из продуктов метаболизма углеводов.