

Возбуждающие химические синапсы
Обратимся к возбуждающим химическим синапсам на спинальных мотонейронах кошки. Эти нейроны имеют шаровидную сому (D = 70 мкм), от которой отходит множество конических дендритов и один аксон. Поверхность перикариона составляет примерно 5- 104 см2, Rм = 600-2000 Ом • см2; См≈6мкФ/см2; т=1,2-5,1 мс. Сома и дендриты густо покрыты синаптическими бутонами и отростками глиальных клеток. Нервные волокна, подходящие к мотонейрону, на расстоянии в 100-200 мкм от него теряют миелиновую оболочку и истончаются (до 0,5- 2 мкм в диаметре). Синаптические бутоны содержат везикулы. Синаптические щели имеют ширину 200 А° (20 нм).
МПП мотонейрона -60------80 мВ. Возбуждающий постсинаптический потенциал, возникающий в соме при приходе одиночного залпа импульсов с афферентов группы 1а (моносинаптической рефлекторной дуги), имеет вид деполяризации с временем возрастания (tв) равным 1,5-2 мс и постоянной времени спада (т) равной 4,7 мс. Амплитуда такого ВПСП, возникающего под одиночным синаптическим входом (при активации одного афферентного волокна), невелика (0,12-0,24 мВ) и стандартна, не зависит от силы раздражения волокна. Но если раздражается многоволоконный задний корешок или периферический нерв, то ответный ВПСП больше и его амплитуда увеличивается с усилением раздражения. Последнее объясняется увеличением количества синхронно активных синаптиче-ских входов на данном нейроне, т. е. количества синхронно возникающих элементарных ВПСП. Причины роста амплитуды здесь, по существу, те же, что и в случае роста ПД нервного ствола . Заметим, что ВПСП одиночных синаптических входов имеют очень низкий квантовый состав (1-2), амплитуды МВПСП соизмеримы с ними (0,12-0,24 мВ).
ВПСП мотонейрона определяются трансмембранным ионным током, по времени соответствующим восходящей фазе ВПСП, возникающим из-за того, что медиатор афферентов (глутамат или субстанция Р?) открывает ионные каналы в постсинаптиче-ской мембране. Характер этого трансмембранного ионного тока был определен в опытах с электрофоретическими инъекциями различных ионов в мотонейрон через микроэлектроды (у кошки и лягушки) и в экспериментах с вариациями межклеточной ионной среды (у лягушки). Оказалось, что ток, порождающий ВПСП, и соответствующий ток в клампе (ВПСТ) -это пассивный, т. е. текущий по электрохимическому градиенту натриевый ток, слабо шунтируемый калиевым током. Е реверсии ВПСП составляет +3--+5 мВ. Падение Rвх (рост проводимости) в момент развития ВПСП (ВПСТ) невелико (∼5%).
ВПСП соседних синаптических входов
суммируются между собой. Так же суммируются
и последовательно возникающие ВПСП.
Эти феномены лежат в основе так называемой
пространственной и временной суммации
возбуждения в ЦНС. Когда общая деполяризация
в соме достигает определенной величины,
возникает ПД нейрона. Здесь, однако,
имеется одна особенность. Дело в том,
что аксонный холмик (начальный сегмент
аксона) относительно сомы по ряду причин
имеет, приблизительно, в 3 раза более
низкий порог электрического раздражения.
Ток, порождаемый ВПСП, выходит через
все внесинаптические участки мембраны
нейрона, но в этих условиях именно в
аксонном холмике он порождает ПД.
Говорят, что холмик играет роль триггера
- спускового крючка. Отсюда ПД
распространяется в аксон, а также
ретроградно в сому. Последнее, видимо,
необходимо для согласования аксонального
и соматического метаболизма. На
электрограмме, записываемой внутриклеточным
электродом, введенным в сому, ПД начального
сегмента и ПД сома-дендритного комплекса
слиты в общий ПД, в котором все же можно
различить эти компоненты. Ступенька на
переднем фронте этого ПД - по с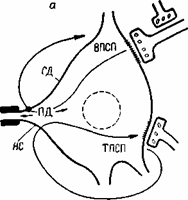 уществу,
ПД начального сегмента, сниженный
расстоянием (λ в сома-дендритном комплексе
составляет около 400 мкм). Отметим одну
особенность ПД сомы мотонейрона. У этого
ПД те же компоненты, что и у аксонального
ПД, но гораздо более сильный следовой
положительный потенциал.
уществу,
ПД начального сегмента, сниженный
расстоянием (λ в сома-дендритном комплексе
составляет около 400 мкм). Отметим одну
особенность ПД сомы мотонейрона. У этого
ПД те же компоненты, что и у аксонального
ПД, но гораздо более сильный следовой
положительный потенциал.
Описанные электрические проявления
синаптической активации характерны
для мотонейронов спинного мозга и других
моторных нейронов ЦНС позвоночных. Но
в интеронейронах картина несколько
иная. Например, в клетках Реншоу в ответ
на одиночный возбуждающий синаптический
залп регистрируется очень длительный
ВПСП, порождающий длинную серию ПД. мпп
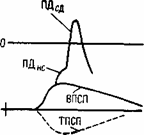
Рис.6.7. Воздействие электрических токов, порождаемых возбуждающим и тормозящим постсинаптическими потенциалами, на выходной участок спинномозгового мотонейрона (начальный сегмент аксона).
а - схема петель токов ВПСП и ТПСП. НС - начальный сегмент аксона; СД - сома-дендритный комплекс. Точка - место выхода петли тока ВПСП; б - схема наложения записей ВПСП, ПДНС и ПДСД. Пунктиром показан ТПСП, который может быть совмещен во времени с ВПСП, если тормозной залп будет послан в спинной мозг несколько раньше возбуждающего.
По-видимому, в синапсах этих клеток имеются условия для существенного продления действия медиатора, а внесинаптиче-ская мембрана этих нейронов обладает очень низкой аккомодационной способностью. В клетках Реншоу триггерной зоной является не аксонный холмик, а соматическая мембрана, прилежащая к синаптическим районам. Их очень краткий ПД (t пика=0,5-1,0 мс) не имеет сильного следового положительного потенциала. Такими свойствами, вероятно, обладают многие вставочные нейроны (интернейроны) ЦНС.
Среди нейронов ЦНС у разных животных (а также среди элементов сетчатки глаза) имеются и такие, возбуждение которых исчерпывается постсинаптическим электрогенезом. Эти клетки не генерируют ПД. У них очень короткие аксоны и поэтому их ВПСП, возникающие в соме, могут чисто электротонически распространяться до окончания аксона, побуждая его к секреции медиатора.