

Лекция 3. Мембранные потенциалы нервных элементов в покое и при возбуждении.
3.1. Мембранный потенциал покоя
У всех исследованных физиологами клеток животных и растений поверхностная мембрана в покое электрически поляризована. В этом можно убедиться, введя внутрь клетки микроэлектрод, соединенный с электрорегистрирующей установкой. Как только кончик микроэлектрода попадает внутрь клетки, он мгновенно (скачком) приобретает некоторый постоянный электроотрицательный потенциал по отношению к электроду, расположенному в окружающей клетку жидкости Величина разности этих потенциалов у нервных клеток и волокон, например гигантских нервных волокон кальмара, составляет около - 70 мВ. Эту величину называют мембранным потенциалом покоя (МПП). Во всех точках аксоплазмы этот потенциал практически одинаков. Если кончик микроэлектрода выводят из клетки возвратным движением или прокалывая клетку насквозь, то разность потенциалов между электродами скачкообразно исчезает. Если кончик микроэлектрода недостаточно тонок, то он входит и выходит из клетки постепенно, давит и тянет за собой ткань, повреждая клетку, это "смазывает" броски потенциала и снижает величину МПП .Заметим, что здесь и дальше снижением мембранного потенциала мы будем называть уменьшение внутриклеточной негативности, вообще смещение внутриклеточного потенциала в позитивную сторону.
Связь МПП с поляризацией мембраны клетки доказывается, например, классическим опытом Бейкера, Ходжкина и Шоу с удалением аксоплазмы из гигантского аксона кальмара (диаметр аксона около 1 мм). Лишенный аксоплазмы аксон при его перфузии солевым раствором, сходным по ионному составу с аксоплазмой, демонстрирует примерно такие же скачки потенциала, т. е. такой же МПП, как и нормальный аксон.
Природа поляризации клеточной мембраны сегодня в основном определена. Чтобы ее понять, необходимо рассмотреть некоторые свойства самой мембраны, а также ионный состав внеклеточной среды и внутриклеточной жидкости. Мембрана аксона кальмара (и всех других клеток)-это очень тонкая, но достаточно прочная оболочка. Согласно электронно-микроскопическим данным ее толщина 5-10 нм (50-100 А). Известно, что мембрана состоит из липидов, белков и мукополисахаридов. Бимолекулярный слой липидов составляет матрикс мембраны. Белки, вкрапленные в липидный матрикс, образуют каналы для воды и ионов, формируют ионные насосы и т. п. (рис.3.1).
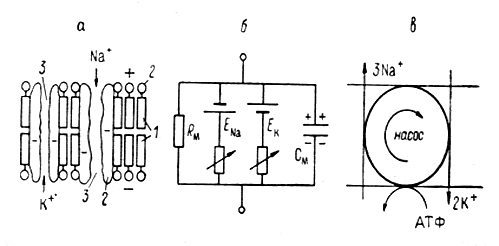
Рис.3. 1. Упрощенные схемы строения плазматической мембраны и ее механизмов, ответственных за формирование потенциала покоя и потенциала действия.
а - схема строения и б - эквивалентная электрическая схема мембран; в - схема работы ионного насоса.
1 - билипидный матрикс, 2- белковые молекулы; 3 - открытые катионные каналы.
Мукополисахариды, располагаясь на поверхности мембраны, осуществляют рецепторные функции. Заметим, что мембрана постепенно расходуется клеткой (при экзоцитозе и эндоцитозе) и постоянно обновляется.
При этом ее качества могут несколько меняться в зависимости от изменения программы соответствующих синтетических процессов.
Мембрана обладает значительными электрическим сопротивлением и емкостью. У аксона кальмара сопротивление мембраны (RM) составляет в покое ∼ 1000 Ом • см2, а емкость (См) - 1 мкФ/см2. Емкость мембраны, которая задается в основном ее липидным матриксом, довольно постоянна. Сопротивление же мембраны проходящему току сильно зависит от состояния ее ионных каналов.
Мембрана проницаема для жирорастворимых веществ, молекулы которых проникают через липидный матрикс. Крупные водорастворимые молекулы, в том числе анионы органических кислот, по существу, совсем не проходят через мембрану (они могут покидать клетку лишь путем экзоцитоза). В то же время в мембране нервного волокна имеются каналы, проницаемые для воды, для молекул водорастворимых веществ и для ионов.
Особый интерес для электрофизиологии представляют каналы для ионов. Среди них имеются неспецифические каналы ионной "утечки", каждый из которых проницаем для К, Na+ и С1-(больше всего для К+). Эти каналы не имеют воротных механизмов, они всегда открыты и почти не меняют своего состояния при электрических воздействиях на мембрану. Кроме того, в мембране нервного волокна присутствуют специальные натриевые и калиевые каналы, т. е. каналы, избирательно (селективно) пропускающие только Na+ или только К+- Эти каналы обладают воротными механизмами и могут быть открытыми или закрытыми. В покое у нервных волокон практически все натриевые каналы мембраны закрыты, а большое число калиевых открыто.
Определенное состояние ионных каналов мембраны (закрытое у натриевых, открытое у значительной части калиевых) очень важно для генерации МПП нервного волокна. Вторым важным для формирования МПП условием является различие ионных составов аксоплазмы и внешней среды, создаваемое непрерывной работой ионных насосов мембраны .
Таблица 3.1.
Ионный состав (мМ на кг воды) аксоплазмы и внешней среды гигантского аксона кальмара |
|||
Ионы |
Аксоплазма |
Кровь |
Морская вода |
K๋+ |
400 |
20 |
10 |
Na๋+ |
50 |
440 |
460 |
С1- |
401-150 |
560 |
540 |
Са2+ |
0,4 |
10 |
10 |
Mg2+ |
10 |
54 |
53 |
Изотионовая кислота |
250 |
- |
- |
Аспарагиновая кислота |
75 |
- |
- |
Глутаминовая кислота |
12 |
- |
- |
Янтарная и фумаровая кислоты |
17 |
- |
- |
АТФ |
0,7 |
1,7 |
|
Аргининфосфат |
1,3-5,7 |
|
|
Н20 |
865 |
870 |
966 |
|
|
|
|
В таблице 3.1 сопоставлены ионные составы аксоплазмы гигантского аксона и крови у кальмара. Различия в этих составах постоянны и в основном сводятся к тому, что в аксоплазме по сравнению с кровью меньше Na+, больше К+ и несравненно больше органических анионов Последние, как уже говорилось, не могут просочиться через неповрежденную мембрану наружу. Что касается катионных различий, то они являются результатом работы так называемого натрий-калиевого насоса мембраны, непрерывно откачивающего Na+ из клетки в обмен на К+ (с использованием энергии АТФ). Заметим, что такой перенос ионов против градиентов их концентраций называют активным ионным транспортом в отличие от пассивного транспорта ионов, идущего по электрохимическим градиентам.
Поляризация мембраны при открытых калиевых каналах, т. е. при высокой калиевой проницаемости мембраны (Рк) и при наличии большого трансмембранного градиента концентраций К+ объясняется прежде всего тем, что имеется некоторая, хотя и очень небольшая, утечка внутриклеточного К+ в среду. Эта утечка и создает разность электрических потенциалов между средой и аксоплазмой. Заметим, что вход Na+ в клетку или выход из нее органических анионов, что могло бы компенсировать нарушения электронейтральности от потери К+, исключены свойствами покоящейся мембраны. В этой ситуации на мембране создается двойной электрический слой (снаружи катионы, главным образом Na+, внутри - анионы, главным образом органических кислот), препятствующий дальнейшему выходу К+ (рис.3.2).

Рис.3.2. Возникновение электрохимического равновесия на полупроницаемой мембране.
Диффузионное давление (1) в равновесии с противоположной электростатической силой (2), стремящейся удержать вместе ионы с противоположными зарядами (по С.Немечек, 1978).
Перемещение К+ из клетки наружу при отсутствии МП осуществляется по концентрационному градиенту этого иона, совершающему "осмотическую" работу. В силу возникающего мембранного потенциала ионы К+ возвращаются в клетку, т. е. совершается "электрическая работа": Aэл.= QE=nFE, где Q - количество электричества, n - валентность, F - число Фарадея, заряд моля одновалентных ионов (96500 Кл/гр • экв), Е - потенциал. По мере выхода ионов К+ растет Аэл. и несколько падает Аосм, в итоге достигается Е на мембране, при котором Аэл = Аосм.. Для иона К+, т. е. калиевый равновесный потенциал в упрощенной форме:
E=58 lg [K]н/ [K]в (формула Нернста).
Разность между текущим значением МП и Ек называют электрохимическим градиентом для К+. Электрохимический градиент - причина пассивного движения К+ (и любого другого иона) через мембрану в естественных условиях.
МПП гигантского аксона кальмара ( - 70 мВ) близок к его Ек ( - 75 мВ), но не равен ему, ибо МПП здесь формирует утечка и других ионов: Na+, Cl-. При этом поступление С1-в аксон, (Еcl> - 70 мВ) повышает, a Na+ понижает МПП (ENa=+55MB).
Итоговая величина Ем, создаваемого утечкой многих ионов, может быть достаточно точно рассчитана по формуле Гольдмана:
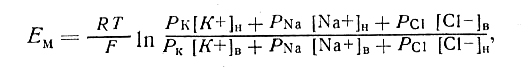
где Р - проницаемость мембраны для соответствующих ионов. Ее часто выражают в относительных величинах, принимая PK+ за 1. Для мембраны аксона кальмара в покое отношение PK: PNa : Рcl= 1 : 0,04 : 0,45
Рассмотренный ионный механизм формирует так называемый концентрационный потенциал (Еконц.) - основную часть реального МПП. В перфузируемом чистым солевым раствором гигантском аксоне кальмара это, по существу, единственный механизм формирования МПП. Но в естественных условиях в образовании МПП участвует еще один, добавочный механизм. Это так называемый прямой электрогенный эффект натрий-калиевого насоса. Напомним, что этот ионный насос работает, потребляя энергию АТФ, и его основным компонентом является мембранная К - Na-АТФаза. В норме АТФ к насосу поступает из аксональных митохондрий. Поэтому в лишенном аксоплазмы перфузируемом аксоне насос работает только при добавлении к перфузату АТФ. Для его работы, кроме того, требуется наличие в среде ионов К+, а внутри волокна - ионов Na+. Дело в том, что макромолекула насоса осуществляет свое рабочее движение - конформацию лишь в случае присоединения к ней снаружи ионов К+, а изнутри клетки - ионов Na+ . Насос не работает в бескалиевых средах и при потере внутриклеточного Na+. Прямой электрогенный эффект насоса (который следует отличать от косвенного, т. е. от участия насоса в создании концентрационных градиентов) состоит в дополнительной поляризации мембраны, получающейся при неравенстве числа (q) ионов Na+ и К+, переносимых в каждом цикле работы насоса. Только если эти числа равны, насос работает электронейтрально.У гигантского аксона кальмара Rм относительно мало и поэтому Енас тоже невелико (1 мВ). В некоторых нервных клетках моллюсков, где Rм велико (мегаомы) - Енлс достигает десятка мВ.
Енас может быть быстро устранен блокадой мембранной Na - К-АТФазы с помощью сердечных гликозидов (уабаин и др.) или за счет снижения температуры до 5° С, наконец, за счет нарушения выработки АТФ (при действии динитрофенола и цианидов). Eконц. при этом не исчезает, а падает достаточно медленно по мере потери ионных градиентов на мембране из-за утечки Na+ и других ионов.
Таким образом, реальный МПП складывается из Еконц и Eнас.
В миелинизированных нервных волокнах у позвоночных потенциал покоя мембраны перехвата Ранвье составляет около 70 мВ. Его концентрационный компонент имеет в основном калиевую природу, как и в аксоне кальмара. Ионы С1- в его формировании принимают незначительное участие. Электрогенный эффект ионного насоса в нормальной среде здесь близок к нулю. И только при повышенной концентрации К+ снаружи [К]н этот эффект усиливается настолько, что может достигнуть 3 - 4 мВ. Последнее достигается за счет резкого усиления насосного тока.
Мембранный потенциал покоя в самой мембране проявляется как электрическое поле значительной напряженности (∼105В/ см). Это поле воздействует на макромолекулы мембраны и придает их заряженным группам определенную пространственную ориентацию. Особенно важно то, что электрическое поле МПП обеспечивает закрытое состояние так называемых активационных ворот натриевых каналов и открытое состояние их инактивационных ворот. Этим обеспечивается состояние покоя и готовности к возбуждению. Даже относительно небольшой сброс мембранного потенциала (частичная деполяризация) открывает активационные ворота этих каналов и выводит клетку из состояния покоя, дает начало возбуждению. При возбуждении, конечно, используется электрическая энергия, накопленная в МПП.
В плазматической мембране нервных и мышечных клеток роль МПП, по-видимому, исчерпывается сказанным. Однако для некоторых других мембран, например внутренней мембраны митохондрии, очевидно участие МПП в энергетических процессах - сопряжении дыхания и фосфорилирования. МПП (∼+200мВ) здесь создается в ходе биологического окисления и связан с возникновением градиента Н+-ионов . Все агенты, переносящие положительные заряды внутрь митохондрии, снижают ее МПП и разобщают дыхание и фосфорилирование.
В заключение остановимся на понятии собственного электрического потенциала мембраны. Под последним подразумевают потенциал поверхностей мембраны относительно ее внутренних частей (∼30м В). Этот потенциал имеет отрицательный знак - на поверхностях и положительный внутри мембраны.
Он определяется наличием выступающих наружу отрицательно заряженных групп (типа СОО-) у многих молекул, формирующих мембрану. Величина этого потенциала определяет способность поверхностей мембраны связывать катионы, в частности. Са2+, что существенно для работы воротных и других ее механизмов.
Собственный потенциал мембраны, поскольку он одинаков у обеих сторон мембраны, не входит в МПП.