

9.3.1. Нисходящие проводящие пути.
Эти пути связывают высшие отделы ЦНС с эффекторными нейронами спинного мозга. Основными из них являются пирамидный, руброспинальный, вестибулоспинальный и ретикулоспинальный тракты.
Пирамидный тракт образован аксонами клеток двигательной зоны коры больших полушарий. Направляясь к продолговатому мозгу, эти аксоны отдают большое число коллатералей структурам промежуточного, среднего, продолговатого мозга и ретикулярной формации. В нижней части продолговатого мозга большая часть волокон пирамидного пути переходит на противоположную сторону (перекрест пирамид), образуя латеральный пирамидный тракт. В спинном мозгу он располагается в боковом столбе. Другая часть волокон идет, не перекрещиваясь, до спинного мозга и только на уровне сегмента, в котором оканчивается, переходит на противоположную сторону. Это прямой пирамидный тракт. Оба они заканчиваются на мотонейронах передних рогов серого вещества спинного мозга. Состав волокон этого пути неоднороден, в нем представлены миелинизированные и немиелинизированные волокна разного диаметра со скоростями проведения возбуждения от 1 до 100 м/с.
Основной функцией пирамидных путей являтся передача импульсов для выполнения произвольных движений. Надежность в осуществлении этой функции повышается благодаря дублированию связи головного мозга со спинным посредством двух путей - перекрещенного и прямого. В эволюционном ряду пирамидный тракт развивался параллельно с развитием коры больших полушарий и достиг наибольшего совершенства у человека.
Руброспинальный тракт (Монакова) образован аксонами клеток красного ядра среднего мозга. Выйдя из ядра, волокна полностью переходят на противоположную сторону. Часть из них направляется в мозжечок и ретикулярную формацию, другие - в спинной мозг. В спинном мозгу волокна располагаются в боковых столбах перед перекрещенным пирамидным путем и оканчиваются на интернейронах соответствующих сегментов. Руброспинальный тракт несет импульсы от мозжечка, ядра вестибулярного нерва, полосатого тела.
Основное назначение руброспинального тракта - управление тонусом мышц и непроизвольной координацией движений. В процессе эволюции этот путь возник рано. Большое значение он имеет у животных, слабее развит у человека.
Вестибулоспинальный тракт образован волокнами, которые являются отростками клеток ядра Дейтерса, лежащего в продолговатом мозгу. Этот тракт имеет наиболее древнее эволюционное происхождение. По нему передаются импульсы от вестибулярного аппарата и мозжечка к мотонейронам вентральных рогов спинного мозга, регулирующие тонус мускулатуры, согласованность движений, равновесие. При нарушении целостности этого пути наблюдаются расстройства координации движений и ориентации в пространстве.
В спинном мозгу помимо основных длинных имеются и короткие нисходящие пути, соединяющие между собой его отдельные сегменты.
Лекция 10. Задний мозг.
10.1. Строение заднего мозга
Задний мозг, состоящий из продолговатого мозга и варолиева моста, представляет собой отдел головного мозга, который является непосредственным продолжением спинного мозга. Серое вещество спинного мозга переходит в серое вещество продолговатого мозга и сохраняет черты сегментарного строения. Однако основная часть серого вещества распределена по всему объему заднего мозга в виде обособленных ядер, разделенных белым веществом.
В продолговатом мозгу симметрично заложены ядра четырех последних пар черепно-мозговых нервов, осуществляющих афферентную и эфферентную иннервацию головы и внутренних органов. В самой нижней части ромбовидной ямки располагается ядро подъязычного нерва (XII пара), иннервирующего мышцы языка. Двигательное ядро добавочного нерва (XI пара) является продолжением заднебокового отдела переднего рога спинного мозга и связано с иннервацией мускулатуры шеи (рис.10.1.).
Довольно поверхностно лежит группа ядер блуждающего нерва (X пара), который является смешанным и одновременно участвует в вегетативной, двигательной и чувствительной иннервации. Соответственно функции блуждающему нерву принадлежат три ядра. Вегетативное ядро дает парасимпатические волокна, идущие к гортани, пищеводу, сердцу, желудку, тонкой кишке, пищеварительным железам. Чувствительные волокна блуждающего нерва от многих внутренних органов в продолговатом мозгу образуют одиночный пучок (солитарный тракт) и заканчиваются в чувствительном ядре солитарного тракта. Третье ядро блуждающего нерва - соматическое, двигательное - лежит глубже в латеральном отделе продолговатого мозга и носит название обоюдного. Нейроны этого ядра рефлекторно регулируют последовательность сокращения мышц глотки и гортани при дыхании.
Языкоглоточный нерв (IX пара), так же как и блуждающий, содержит двигательные, чувствительные и вегетативные волокна. Двигательные волокна участвуют в иннервации мышц ротовой полости и глотки.
Чувствительные волокна от вкусовых почек задней трети языка слизистой оболочки глотки входят вместе с волокнами блуждающего нерва в одиночный пучок. Отростки нейронов, формирующих вегетативное ядро языкоглоточного нерва, идут к парасимпатическим ганглиям, которые связаны с иннервацией секреторного аппарата слюнных желез.
На границе продолговатого мозга и моста располагаются ядра слухового, или вестибулокохлеарного нерва (VIII пара). Этот нерв состоит из двух частей: нерва улитки и нерва преддверия. Волокна нерва улитки, или собственно слухового нерва, представляют собой аксоны клеток лежащего в улитке спирального ганглия. Эти волокна оканчиваются в кохлеарных ядрах (рис. ).
Волокна нерва преддверия формируются аксонами клеток узла преддверия, локализованного на Дне внутреннего слухового прохода. Эти волокна заканчиваются в трех ядрах моста: медиальном ядре Швальбе, латеральном ядре Дейтерса и верхнем ядре Бехтерева. Отростки нейронов второго порядка из этих ядер направляются к фастигиальному ядру мозжечка. Из ядра Дейтерса, которое можно отнести к надсегментарным образованиям, начинается вестибулос-пинальный тракт, идущий в спинной мозг.
В латеральном отделе ретикулярной формации моста расположено ядро лицевого нерва (VII пара). Этот нерв является смешанным. Афферентные чувствительные волокна лицевого нерва передают информацию от вкусовых рецепторов передней части языка. Вегетативные волокна участвуют в иннервации подчелюстных и подъязычных слюнных желез. Эфферентные волокна лицевого нерва имеют концевые разветвления в мимической мускулатуре лица.
Ядро отводящего нерва (VI пара) заложено поверхностно в задней половине моста. Моторные волокна этого нерва иннервируют мышцы, двигающие глазное яблоко, чувствительные волокна связаны с проприоцепторами этих мышц.
И наконец, последняя пара черепно-мозговых нервов, относящаяся к заднему мозгу, - это тройничный нерв (V пара). Этот нерв смешанный и обладает двигательными и чувствительными ядрами. Двигательное ядро расположено в верхнем отделе покрышки моста. Нейроны этого ядра участвуют в иннервации жевательных мышц, мышцы нёбной занавески и мышцы, напрягающей барабанную перепонку. Длинное, простирающееся через весь мост чувствительное ядро тройничного нерва содержит клетки, к которым подходят афферентные аксоны от рецепторов кожи лица, слизистой оболочки носа, зубов, надкостницы костей черепа (рис.10.1.).
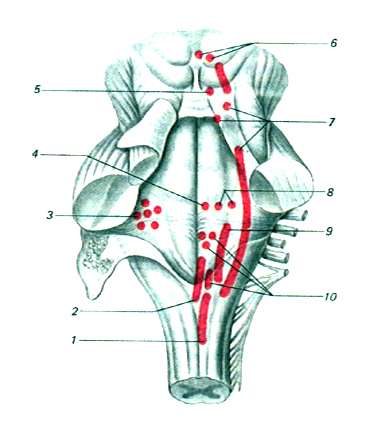
Рис.10.1. Расположение ядер черепно-мозговых нервов в стволовой части мозга:
1 - добавочный нерв, 2 - подъязычный, 3 - слуховой, 4 - отводящий, 5 - блоковый, 6 - глазодвигательный, 7 - тройничный, 8 - лицевой, 9 - языкоглоточный, 10 - блуждающий нерв.
Изучая топографию серого вещества заднего мозга, необходимо отметить наличие в нем ядер чувствительных путей задних столбов спинного мозга. Это ядра тонкого пучка Голля и клиновидного пучка Бурдаха, являющихся путями, которые проводят информацию от тактильных и проприоцепторов, сигнализирующих о положении тела в пространстве. Кроме того, внутри заднего мозга расположено верхнее оливарное ядро, включенное в восходящий путь, который передает акустическую информацию. По латеральной поверхности продолговатого мозга поднимается продолжение боковых столбов спинного мозга.
Центральная часть заднего мозга занята сетевидной, или ретикулярной, формацией, которая представляет собой скопление нервных клеток, напоминающее по принципу организации нервные сети кишечнополостных. Ретикулярные нейроны и распределяются диффузно, и группируются в ядрах, среди которых можно выделить наиболее крупные: ретикулярное гигантоклеточное, ретикулярное мелкоклеточное, латеральное ретикулярное ядро на уровне продолговатого мозга, каудальное ретикулярное и оральное ретикулярное на уровне моста. По цитоархитектоническим особенностям в ретикулярной формации заднего мозга выделяют медиальную часть с множеством нейронов крупного размера и латеральную, включающую в себя более мелкие элементы. По своей топографии ретикулярная формация ствола мозга является структурой, расположенной на пути всех входных и выходных систем мозга.
К дендритам и соме ретикулярных нейронов подходят коллате-рали от аксонов сенсорных восходящих путей, локализованных в вентральной и латеральной частях стволовой части мозга. С другой стороны к нейронам ретикулярной формации подходят коллатерали нисходящих моторных трактов, идущих из переднего мозга. От нейронов гигантоклеточного и каудального ретикулярного ядра начинается нисходящий ретикулоспинальный тракт, устанавливающий прямые связи с мотонейронами спинного мозга. Ретикулярная формация имеет связи с ядрами черепно-мозговых нервов, с мозжечком, промежуточным мозгом и через последний - с корой больших полушарий.
Таким образом, богатство связей ретикулярной формации свидетельствует о том, что она является надсегментарной структурой, приспособленной к регуляции деятельности различных отделов центральной нервной системы.