

Проблема репликации концов линейных молекул
Модель "заячьи уши".
|
|
Работает у ряда вирусов. |
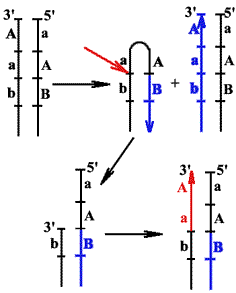
Репликация концов ДНК хромосом эукариот
Удаление РНК-праймеров после завершения синтеза линейных ДНК в виде фрагментов Оказаки и заделывание образующихся между фрагментами брешей нуклеотидами ДНК приводит к тому, что дочерние цепи ДНК оказываются короче материнских на размер первого РНК-праймера (10-20 нукл.).
Образуются 3'-оверхенги, т.е. выступающие 3'-концы материнских цепей. Они узнаются теломеразой - ферментом, содержащим помимо белковой части еще и РНК, выполняющую роль матрицы для наращивания ДНК повторами.
Теломераза последовательно наращивает материнские цепи ДНК повторами, используя 3'-оверхенги в качестве затравок. Образующиеся длинные одноцепочечные концы в свою очередь служат матрицами для синтеза дочерних цепей традиционным репликативным механизмом.
|
|

Хромосомы соматических клеток человека фланкированы многократно повторенными гексамерами TTAГГГ, общая длина районов с повторами может достигать 10 тыс. пар нуклеотидов. В комплексе со специфическими белками такие тандемные повторы образуют теломеры, защищающие концы ДНК от действия экзонуклеаз, предотвращающие неправильную рекомбинацию и позволяющие концам хромосом прикрепляться к ядерной оболочке.
При каждом раунде репликации происходит укорочение теломер в среднем на 50 пар нуклеотидов. Поскольку теломерные последовательности не являются кодирующими, они выступают в роли буферной зоны - как защита от "проблемы концевой репликации".
Укорочение ДНК в ходе каждого раунда репликации лишь сокращает нетранскрибируемый текст теломеры, но не приводит к утрате смысловых последовательностей - генов и регуляторов их экспрессии.
![]()
Регуляция репликации прокариот известна, т.к. известны гены белков регуляции репликации E.сoli и механизмы их включения (выключения). Для эукариот эти механизмы еще не ясны, но известно расписание репликации ДНК по разным хромосомам.
![]()
Причины ошибок при синтезе днк
Способность ошибаться заложена в самой структуре фермента.
![]()
Скорее всего, ферменты, которые не ошибались, были тупиковыми ветвями эволюции. На первых этапах зарождения жизни разнообразие обеспечивалось только такими ошибками.
In vitro происходит 1 ошибка на 100 тыс. Нукл. Для средней днк-полимеразы.
In vitro можно уменьшить вероятность ошибки до 1 на 1млн. Нукл., если добавить ssb, геликазу и лигазу.
Можно и увеличить вероятность ошибки до 1 на 100, если дать неадекватное количество субстрата, а также если добавить ионы серебра, бериллия, меди, кобальта, никеля, свинца. Это происходит из-за конкуренции этих ионов с ионами магния за связывание с ДНК-полимеразой.
Еще один способ повышения количества ошибок - добавление аналогов нуклеотидов. Например бромдезоксиуридина - аналога тимидина.
![]()
Это одно из средств борьбы со СПИДом и раком. Аналоги одинаково вредны для всех клеток, однако в пораженных вирусом клетках чаще проходит репликация.