

Позитивный контроль работы lac-оперона
Lac-оперон, подчиняющийся схеме негативной индукции, имеет и позитивный контроль.
|
цАМФ образуется из АТФ ферментом аденилатциклазой. Фосфодиэстераза превращает цАМФ в АМФ. Глюкоза активирует второй и инактивирует первый фермент. Чем больше в клетке глюкозы, тем меньше цАМФ. |
|
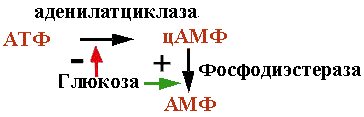
![]()
|
Если нет глюкозы, то цАМФ соединяется с белком катаболической репрессии (САР) и образуется комплекс САР·цАМФ, активирующий посадку РНК-полимеразы на промотор. В присутствии лактозы lac-оперон включается и работает. |
|
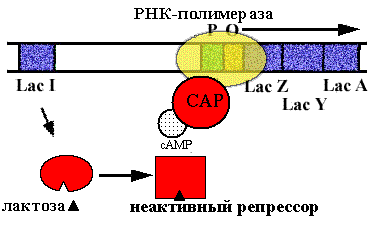
Если же в клетке есть еще и глюкоза (более экономичный источнок энергии), то нет цАМФ - и активатор не образуется, lac-оперон работает "вяло", без дополнительной индукции.
Синтез белка в клетке состоит из двух этапов: рекогниции и собственно синтеза полипептида на рибосоме. Ключевым субстратом рекогниции является транспортная РНК.
Структура транспортной рнк
Транспортные РНК (tРНК) - короткие молекулы (70-90 нукл.), имеющие и вторичную, и третичную структуру.
|
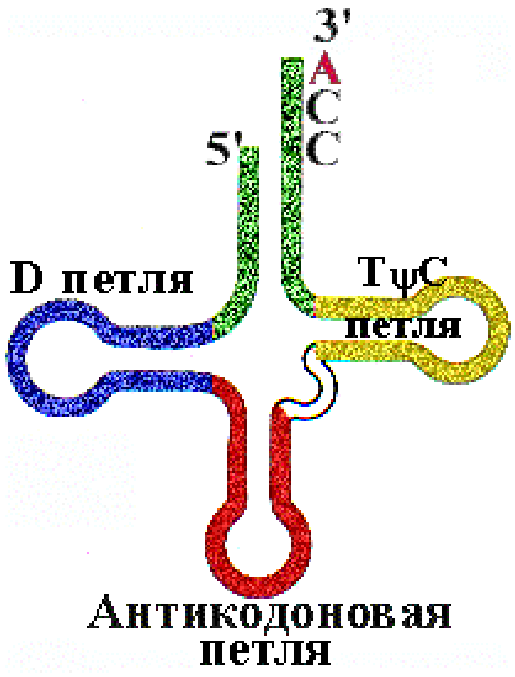
Вторичная структура - "клеверный лист". Последовательность CCA на 3'-конце одинакова для всех tРНК. К концевому аденозину (А) присоединяется аминокислота. Наличие в tРНК тимина (T), псевдоуридина( ) (в TC- петле ), и дигидроуридина (ДГУ) (в D-петле) - минорных, т.е. редко встречающихся в РНК нуклеотидов, указывает на особенности ее строения, необходимые для безошибочного узнавания ферментами, для защиты от действия рибонуклеаз (поэтому tРНК - долгоживущие, в отличие от mРНК). |
|
|
|
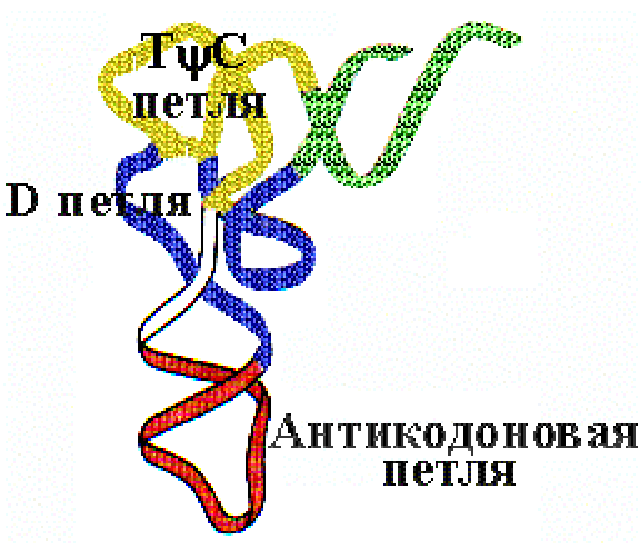
Третичная структура в проекции на плоскость имеет форму бумеранга. Разнообразие первичных структур tРНК - 61+1 - по количеству кодонов (соответственно числу антикодонов в tРНК) + формилметиониновая tРНК, у которой антикодон такой же, как у метиониновой tРНК. Разнообразие третичных структур - 20 (по количеству аминокислот). |
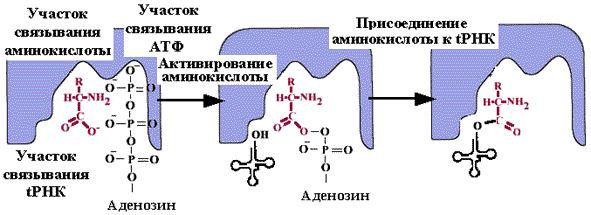
Рекогниция
Определение: рекогниция - это подготовительный этап трансляции, суть которого в образовании ковалентной связи между tРНК и соответствующей аминокислотой.
![]()
Состоит из двух стадий:
1. Активирование аминокислоты.
2. Присоединение аминокислоты к tРнк - аминоацилирование.
Обе стадии рекогниции осуществляются ферментом аминоацил-tРНК-синтетазой (APC-азой, кодазой). Существует 20 вариантов кодаз (по числу аминокислот). У каждой кодазы 3 центра опознавания. Каждая АРС-аза узнает третичную структуру tРНК.
|
|
![]()
Определение: tРНК, имеющие разную первичную, но одинаковую третичную структуру, акцептируют одну и ту же аминокислоту и называются изоакцепторными tРНК.
![]()
|
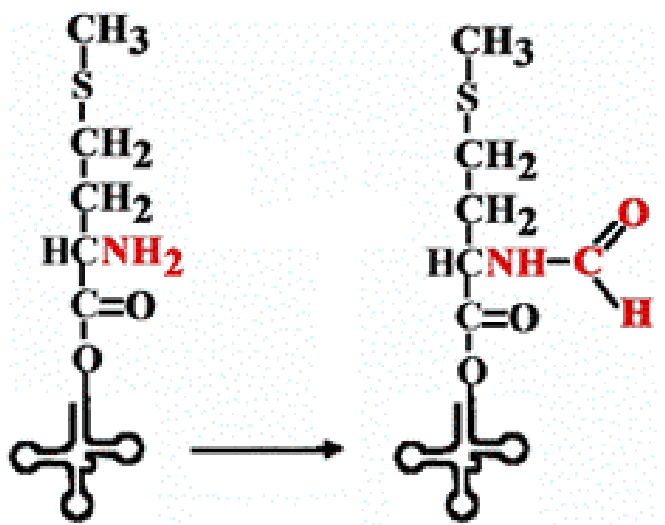
Есть особая tРНК, которая называется формилметиониновой tРНК. Она узнается метиониновой кодазой, соединяется с метионином и уже после реакции аминоацилирования метионин формилируется специальным ферментом, который узнает эту особую форму tРНК. Именно с формилметионина начинается синтез любого полипептида у прокариот. |
|
![]()
Определение: аминоацилирование - это образование связи между аминокислотой и tPHК.
Следующий этап трансляции - собственно синтез полипептидов, происходит на рибосомах.