

1974 Г. Оказаки.
![]()
Рифампицин - ингибитор бактериальной РНК-полимеразы (на стадии инициации).
Хлорамфеникол - ингибитор трансляции на бактериальных рибосомах.
![]()
Если одновременно с заражением E. сoli фагом добавить хлорамфеникол, то блокируются трансляция, репликация II и сборка фагов.
Если подействовать рифампицином, то блокируется не только транскрипция и все следующие процессы, но и репликация I.
![]()
Вывод: бактериальная РНК-полимераза участвует в репликации ДНК фага.
Вся фаговая ДНК составляет ~6000 нукл.
![]()
Определение: origin (ori) - район начала репликации.
![]()
|
|
В районе ori (начало репликации) имеется 4 шпильки. Эти шпильки опознаются РНК-полимеразой, и вторая шпилька используется в качестве матрицы. По мере образования РНК шпилька плавится. Образуется РНК-затравка длиной 24 нукл., 3'-конец которой используется ДНК полимеразой III. |

|
Когда
3'-конец синтезируемой цепи ДНК
"утыкается" в 5'-конец РНК-затравки,
ДНК-полимераза III вытесняется
ДНК-полимеразой I, которая, обладая
5' |
|
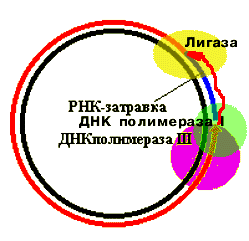
Все эти ферменты (ДНК-полимераза III, ДНК-полимераза I, лигаза) входят в состав реплисомы. Они представляют единый белковый комплекс, который реагирует изменением конформации на выполнение очередной функции.
Протяженная
(более 100 нукл.) одноцепочечная ДНК-матрица
может быть использована ДНК полимеразой
III, когда полимераза III представлена в
форме holo-фермента. Помимо субъединицы
![]() ,
обладающей полимеразной активностью,
в holo-фермент входит еще несколько
субъединиц, обеспечивающих высокую
процессивность синтеза ДНК.
,
обладающей полимеразной активностью,
в holo-фермент входит еще несколько
субъединиц, обеспечивающих высокую
процессивность синтеза ДНК.
Фаг
![]() X174
X174
Репликация ДНК этого фага не зависит от рифампицина.
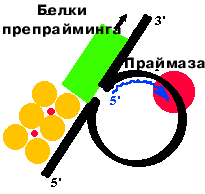
Здесь работает не обычная РНК-полимераза, а особый фермент - праймаза. Он умеет делать только РНК-затравку.
Праймаза нуждается в дополнительных факторах - белках препрайминга. Точно так же (с помощью праймазы и белков препрайминга) образуются РНК-затравки при репликации ДНК E. сoli.
|
|
Праймосома - это белки препрайминга и праймаза. Белки
движутся по матричной цепи ДНК в 5'
|
Топологические проблемы репликации днк
SSB
Белки Альбертса,
обнаруженные в 1968г., снижают температуру
плавления ДНК in vitro на 20-40![]() С.
Они связываются с ДНК электростатически,
хотя имеют отрицательный заряд. Эти
белки содержат кластер положительно
заряженных аминокислотных остатков,
но общий заряд белка отрицателен. У
них повышенное сродство к одноцепочечной
ДНК. Белок не
связывается с двуцепочечной ДНК, не
имеющей расплавленных участков.
С.
Они связываются с ДНК электростатически,
хотя имеют отрицательный заряд. Эти
белки содержат кластер положительно
заряженных аминокислотных остатков,
но общий заряд белка отрицателен. У
них повышенное сродство к одноцепочечной
ДНК. Белок не
связывается с двуцепочечной ДНК, не
имеющей расплавленных участков.
Но если есть одноцепочечная ДНК, то белки легко садятся на нее, выпрямляют ее, превращая ДНК в "палку".
Белки связываются с двуцепочечной ДНК, если в ней есть нарушения вторичной структуры.
|
|
Когда в ДНК образуется расплавленный участок, белок покрывает его за счет электростатических взаимодействий. При этом проявляется сродство белков друг к другу. Они покрывают ДНК сплошным слоем (стехиометрическое количество белка). |

Белки, сидящие на комплементарных цепях, не дают цепям схлопнуться, т.к. имеют мощный отрицательный заряд. Называются эти белки SSB (single strand bind).
![]()
Они не денатурируют ДНК, а лишь фиксируют одноцепочечное состояние.
![]()
Участие SSB в репликации абсолютно необходимо. Они удерживают матричные цепи ДНК в репликативной вилке в одноцепочечном состоянии, а также защищают одноцепочечную ДНК от действия нуклеаз. Они избирательно стимулируют работу ДНК-полимеразы. РНК-полимераза не может использовать одноцепочечную ДНК, покрытую SSB. Избирательность касается и вида. Например, SSB фага Т4 стимулирует ДНК- полимеразу фага Т4, но не ДНК-полимеразы Е. сoli.
![]()
SSB не ферменты - они нужны в стехиометрическом количестве.
![]()