

Регуляция образования рибосомных рнк и белков рибосом e.Сoli
Ежеминутно в E.сoli образуется около 500 рибосом (всего в клетке 10000-50000 рибосом).
Имеется 7 оперонов, в которых закодированы rРНК (всего 3 разных rРНК х 7оперонов = 21 ген). В формировании рибосом участвуют 52 различных белка, а значит 52 гена, их кодирующих. В итоге,
73 Гена должны работать координированно, чтобы не было избытка белков или rРнк.
![]()
Вначале образуется про-rРНК, которая метилируется и процессируется (т.е. "созревает").
|
|

Количество rРНК регулируется количеством рибосомных оперонов, скоростью их транскрипции и работой ферментов метилаз и эндонуклеаз.
![]()
Имеется 7 разных оперонов, в которых закодированы рибосомные белки. Регуляция каждого из них осуществляется отдельно.
-оперон регулируется белком S4.
|
|
Если в клетке имеется свободная 16S rРНК, то S4 связывется с ней (1). Если же 16S rРНК не хватает, то он связывается с mРНК, считывающейся с данного оперона (2). Причем связывается в районе лидера и тем самым мешает трансляции. |
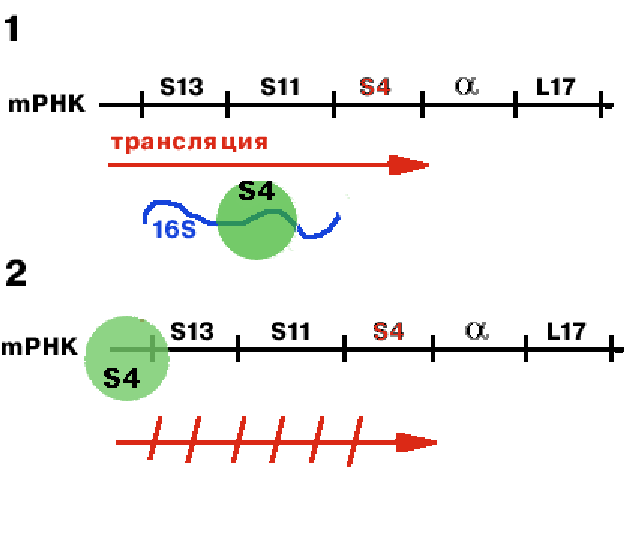
Таким образом, осуществляется регуляция на уровне трансляции.
![]()
Оперон S10 регулируется белком L4.
|
|
РНК-полимераза синтезирует первую лидерную последовательность, длиной 140 нукл. Если 23S rРНК не хватает (2), то белку L4 не с чем соединяться, и он взаимодействует с лидерной последовательностью, придавая ей такую конформацию, которая не позволяет РНК-полимеразе продолжать транскрипцию. В результате синтез mРНК обрывается на первом же лидере (2). |
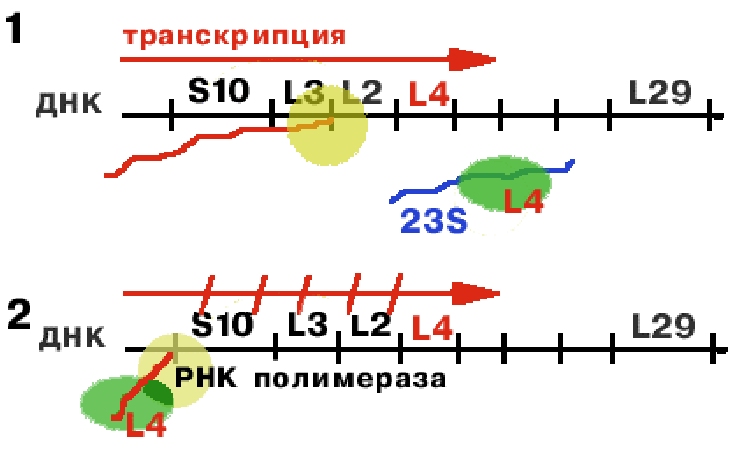
Регуляция на уровне транскрипции.
![]()
Оперон .
|
|

В этом опероне закодированы белки, имеющие принципиальное значение для инициации транскрипции (-фактор), инициации репликации (dna G - праймаза) и инициации трансляции (белок S21). Каждый белок нужен в разном количестве. S21 ~ 50000 копий, dnaG ~ 50, - фактор ~ 5000. Между геном S21 и геном dnaG есть слабый терминатор транскрипции. Ген dnaG имеет инициирующий кодон ГУГ (а не АУГ), который гораздо хуже узнается рибосомой и реже, чем АУГ инициирует трансляцию.
![]()
Транскрипция у эукариот
У эукариот процессы транскрипции и трансляции разобщены во времени и пространстве (транскрипция - в ядре, трансляция - в цитоплазме).
![]()
У эукариот существуют специализированные РНК-полимеразы.
В ядре выделяют 3 типа РНК-полимераз:
РНК-полимераза I - синтезирует rРНК (кроме 5S rРНК).
РНК-полимераза II - синтезирует mРНК и некоторые sРНК.
РНК-полимераза III - синтезирует tРНК, некоторые sРНК и 5SrРНК.
![]()
РНК-полимеразы различаются количеством субъединиц, их аминокислотным составом, и зависимостью от катионов магния и марганца. Для РНК-полимераз I и III необходимое для работы соотношение [Mn2+]/[Mg2+] = 2. Для РНК-полимеразы II - [Mn2+]/[Mg2+] = 5.
Наиболее яркое
различие - чувствительность к
![]() -
аманитину (токсину бледной поганки). Он
полностью подавляет работу РНК-полимеразы
II в концентрации 10-8 М и РНК-полимеразы
III ( в концентрации 10-6 М). РНК-полимераза
I фактически нечувствительна к этому
токсину.
-
аманитину (токсину бледной поганки). Он
полностью подавляет работу РНК-полимеразы
II в концентрации 10-8 М и РНК-полимеразы
III ( в концентрации 10-6 М). РНК-полимераза
I фактически нечувствительна к этому
токсину.
Помимо ядерных РНК-полимераз у эукариот есть еще РНК-полимеразы хлоропластов и митохондрий. Они кодируются в ядре, а не в соответствующих органеллах.
В органеллах образуются свои tРНК, rРНК и рибосомные белки.