

Доказательство полуконсервативного характера репликации
Для выяснения вопроса о характере расхождения цепей по дочерним молекулам Мэтт Мезельсон и Фрэнк Сталь в 1958г. разработали метод равновесного центрифугирования в градиенте плотности CsCl. .
ДНК разделяется не по молекулярным весам, а по удельной плотности.
E. сoli выращивали на протяжении нескольких поколений на среде, содержащей тяжелый изотоп азота (N15), для того, чтобы вся ДНК была "тяжелой". Перед очередным раундом деления клетки синхронизировали. При этом в среде заменяли N15 на легкий изотоп N14 с тем, чтобы вновь синтезированные цепи были "легкими". После репликации ДНК выделяли и центрифугировали в градиенте плотности CsCl.
![]()
Полуконсервативность означает, что каждая дочерняя ДНК состоит из одной матричной цепи и одной вновь синтезированной.
![]()
|
|

Равное распределение "тяжелых" и "легких" цепей между всеми молекулами исключало возможность консервативного способа, согласно которому одна дочерняя клетка получает материнскую ДНК, а другая - вновь синтезированную, обе цепи которой являются новыми.
Клетки второго поколения содержали как полностью "легкие" молекулы, так и "гибридные", состоящие из одной "легкой" и одной "тяжелой" цепи, аналогичные молекулам первого поколения. Этот факт исключал возможность дисперсного механизма, согластно которому куски материнской ДНК случайным образом распределяются между дочерними молекулами.

Понятие о матрице и затравке
Продукты, образуемые в ферментативной системе in vitro.
|
|
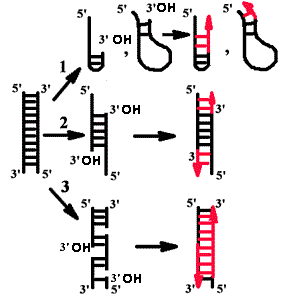
Во всех случаях матрицей для синтеза новых цепей служит одноцепочечная ДНК. Затравкой является 3'-гидроксильный конец двуцепочечной ДНК, причем он должен быть спарен с матрицей.
В том случае, если эндонуклеаза вносила ники с 3'-фосфатным концом, ДНК не являлась активированной.
![]()
Прямым доказательством того, что затравка - 3'-гидроксильный конец, является эксперимент с дидезоксинуклеозидтрифосфатом.
|
|
Если такой активированный нуклеотид сделать меченым по α-фосфату, то он включается в растущую полимерную цепь и всегда обнаруживается на ее 3'-конце. Это говорит о том, что он сам включается, но дальнейший рост цепи невозможен, т.к. нет 3'-гидроксильного конца. |

![]()
Это
также доказывает и униполярность
репликации в направлении 5'
![]() 3'.
3'.

Схема непрерывной антипараллельной репликации in vivo по Корнбергу
1960Г. Гипотетическая модель.
|
|
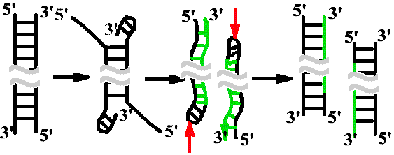
Суть предположения: неизвестный фактор денатурирует концы линейной молекулы, 3'-ОН-концы загибаются и служат затравками для работы ДНК-полимеразы. Фермент осуществляет денатурацию матричной ДНК по мере продвижения и синтеза дочерних цепей. На выходе - дочерние молекулы, которые короче на загнутый конец, т.к. эндонуклеаза вносит разрыв в материнскую цепь.
Схема непрерывной параллельной репликации Джона Кэрнса
|
|

|
1963г. Авторадиография |
|

|
Бактерии выращивались на среде с радиоактивным H3-тимидином в течение 5-и, 10-и, 15-и мин., после чего проводили авторадиографию. |
|
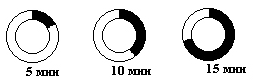
Ширина полос засветки соответствовала двум меченым нитям ДНК (если помечена одна цепь, то полоса будет получаться уже).
В модели допускалось
наличие фермента, способного вести
синтез в направлении 3'![]() 5'. Такой фермент не найден и сегодня.
5'. Такой фермент не найден и сегодня.
![]()
Модель неверна, однако, она побудила искать новые ДНК-полимеразы и в Е. coli были найдены еще две: II и III.
![]()