

Проприоцептивный анализатор
Проприоцептивный анализаторспециализируется навосприятии информации о состоянии опорно-двигательного аппаратаикосвенно участвует в координации движений. Рецепторная часть этого анализатора заложена в опорно-двигательном аппарате и представлена дендритами чувствительных нейронов, оплетающими интрафузальные мышечные волокна в скелетных мышцах, либо многократно разветвляющимися между коллагеновыми или эластическими волокнами связок и сухожилий, либо входящими в состав телец Фатера-Пачини в капсулах суставов.Проприорецепторы обеспечивают восприятие изменения длины и скорости сокращения скелетных мышц (мышечные веретена), степени растяжения сухожилий (тельца Гольджи), а также изменения давления в капсулах суставов (тельца Фатера-Пачини). Информация от рецепторов опорно-двигательного аппарата туловища и конечностей проходит через спинальные ганглии в спинной мозг, а от опорно-двигательного аппарата головы и шеи – через гассеров узел тройничного нерва, коленчатый узел лицевого нерва и каменистый узел языкоглоточного нерва в головной мозг (к чувствительным ядрам этих нервов, заложенным в ромбовидной ямке). Аксоны нейронов спинальных ганглиев, проводящие проприоцептивную информацию в направлении коры больших полушарий, следуют через спинной мозг транзитом в составе путей Голя и Бурдаха и достигают продолговатого мозга, где переключаются на нейроны ядер Голя и Бурдаха. Аксоны же нейронов этих ядер вместе с волокнами чувствительных ядер черепномозговых нервов, имеющих отношение к проведению проприоцептивной информации от головы и шеи, восходят кталамусу(латеральная специфическая группа ядер и вентральная неспецифическая группа ядер) в составемедиальной петли. Нейроны таламических бугров посылают свои проекции к корковому представительству проприоцептивного анализатора, заложенному в теменной доле больших полушарий (постцентральная извилина и верхняя теменная долька), часть волокон таламических нейронов направляется непосредственно к моторной коре (пердцентральная извилина и околоцентральная долька).
Периферическая часть проприоцептивного анализатора заложена в опрно-двигательном аппарате (секлетные мышцы, связки, сухожилия, капсулы суставов). Несмотря на существующее разнообразие рецепторов опрно-двигательного аппарата все они с функциональной точки зрения являютсяпервичночувствующими механорецепторами.
Рецепторы скелетных мышц – мышечные веретена (реагируют собственно на изменение длины мышцы, а также на скорость изменения ее длины)
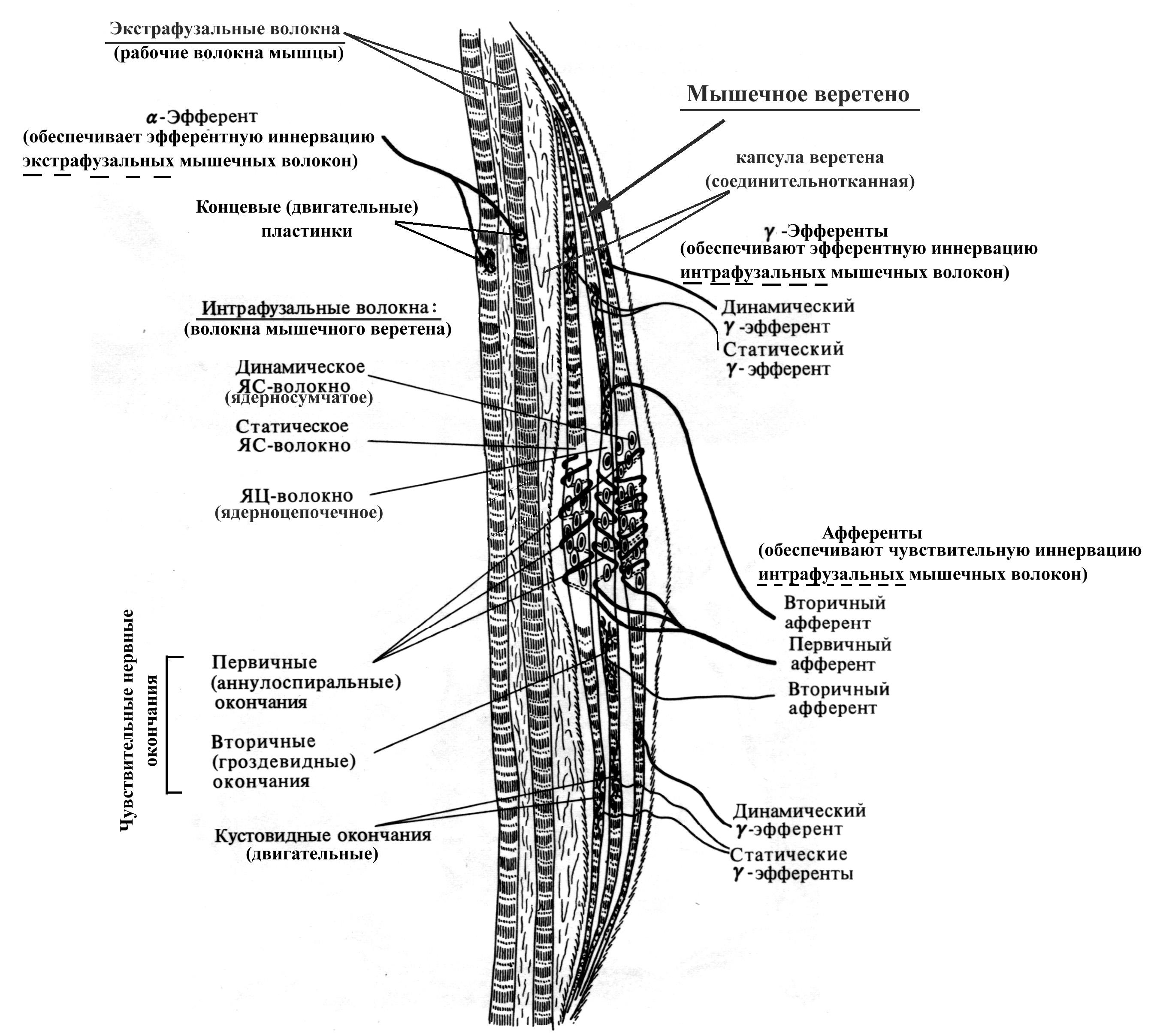
Рис. 16. Схема строения мышечного веретена
Длина мышечных веретен составляет около 3-5 мм, толщина 0,2 мм, ориентированы в мышцах продольно (в связи с этим при растяжении мышцы растягивается и веретено). Каждое веретено содержит от 2 до 12 интрафузальных мышечных волокон двух типов:
ядерно-сумчатые (в состав веретена входит от 1 до 4 таких волокон, толще и длиннее ядерно-цепочечных, ядра образуют скопления в центре волокна), получают афферентную иннервацию только из одного источника (т.н. первичный афферент, спирально закручиваясь, оплетает ядерную сумку);
ядерно-цепочечные (в составе веретена насчитывается до 10 таких волокон, тоньше и короче ядерно-сумчатых, ядра расположены под сарколеммой в виде цепочки на всем протяжении волокна), получают афферентную иннервацию из двух источников – первичный афферент (более толстый, чем вторичный) оплетает ядерно-цепочечное волокно по центру, а вторичный афферент оплетает ядено-цепочечное волокно по одному с каждой стороны от нервных окончаний первичного афферента.
Интрафузальные мышечные волокна ориентированы параллельно экстрафузальным, в связи с чем при растяжении мышцы нагрузка на веретена увеличивается, а при сокращении – уменьшается. Адекватным раздражителем для чувствительных нервных волокон, оплетающих интрафузальные мышечные волокна, является растяжение мышцы, тогда как ее укорочение деактивирует афференты мышечных веретен.
В расслабленной мышце афферентная импульсация от веретен очень слабая, тогда как при растяжении мышцы она резко возрастает. Причем первичные афференты, оплетающие интрафузальные мышечные волокна, реагируют на степень и на скорость растяжения мышцы (динамический ответ), а вторичные, оплетающие ядерно-цепочечные интрафузальные волокна, – только на степень растяжения мышцы (статический ответ).
Наряду с афферентной иннервацией оба типа волокон мышечных веретен получают эфферентную иннервацию со стороны -мотонейронов. Физиологическая роль эфферентной иннервации интрафузальных мышечных волокон состоит в инициации сокращения их периферических частей, и как следствие, усилении натяжения, а значит и повышении чувствительности на растяжение при любой данной длине мышцы. Кроме того, постоянная базальная активность -мотонейронов способствует поддержанию в постоянном тонусе интрафузальных мышечных волокон, что обеспечивает постоянное возбуждение афферентов мышечных веретен и как следствие, -мотонейронов, получающих информацию от этих афферентов и иннервирующих экстрафузальные (рабочие) волокна мышцы. Тоническая же активность -мотонейронов сопровождается поддержанием постоянного (базального) тонуса всей скелетной мышцы. Наряду с описанной зависимостью между возбуждением - и -мотонейронов, имеет место и обратная --коактивация. Причем уровень возбуждения -системы тем выше, чем интенсивнее возбуждены -мотонейроны данной мышцы, т. е, чем больше сила сокращения.
С амо
по себе сокращение мышцы сопровождается
дезактивацией афферентов мышечных
веретен (они активируются в ответ на
растяжение мышцы), но возбуждение-мотонейронов
приводит к сочетанной активации и
-мотонейронов,
вызывающих сокращение интрафузальных
мышечных волокон и, как следствие,
активацию первичных и вторичных
афферентов. Следовательно, состояние
афферентов мышечных веретен в активной
мышце является результатом двух
противоположных факторов: степени
укорочения мышцы (дезактивирующей
афференты веретен) и состояния -системы,
активирующейся сочетанно с возбуждением
-мотонейронов
и вызывающей повышение импульсации в
афферентах мышечных веретен. Во время
активных движений актвированные
-мотонейронами
-мотонейроны,
инициируя сокращение интрафузальных
мышечных волокон, поддерживают постоянную
активность афферентов мышечных веретен
укорачивающейся мышцы, что дает
возможность рецепторам реагировать на
вызванные помехами неравномерности
движения как увеличением, так и уменьшением
частоты им пульсации и участвовать
таким образом в коррекции движений.
амо
по себе сокращение мышцы сопровождается
дезактивацией афферентов мышечных
веретен (они активируются в ответ на
растяжение мышцы), но возбуждение-мотонейронов
приводит к сочетанной активации и
-мотонейронов,
вызывающих сокращение интрафузальных
мышечных волокон и, как следствие,
активацию первичных и вторичных
афферентов. Следовательно, состояние
афферентов мышечных веретен в активной
мышце является результатом двух
противоположных факторов: степени
укорочения мышцы (дезактивирующей
афференты веретен) и состояния -системы,
активирующейся сочетанно с возбуждением
-мотонейронов
и вызывающей повышение импульсации в
афферентах мышечных веретен. Во время
активных движений актвированные
-мотонейронами
-мотонейроны,
инициируя сокращение интрафузальных
мышечных волокон, поддерживают постоянную
активность афферентов мышечных веретен
укорачивающейся мышцы, что дает
возможность рецепторам реагировать на
вызванные помехами неравномерности
движения как увеличением, так и уменьшением
частоты им пульсации и участвовать
таким образом в коррекции движений.
Рецепторы связок и сухожилий – тельца Гольджи(реагируют на растяжение связок и сухожилий)
Имеют строение, подобное тельцам Руффини соединительной ткани кожи, но отличаются большими размерами. Находятся в зоне соединения мышечных волокон с сухожилием и расположены последовательно по отношению к мышечным волокнам.
Сухожильные рецепторы Гольджи слабо реагируют на растяжение мышцы, но возбуждаются при ее сокращении; интенсивность их импульсации пропорциональна силе сокращения мышцы, что позволяет рассматривать их как источник информации о силе, развиваемой мышцей. На спинальном уровне афферентные нервные волокна, идущие от рецепторов Гольджи, вызывают торможение мотонейронов собственной мышцы и возбуждение мотонейронов мышцы-антогониста.
Рецепторы капсул сустав – тельца Фатера-Пачини (рецепторы, реагирующие на изменение давления в капсуле сустава, а следовательно, на его положение или изменение суставного угла). Подобно проприорецепторам скелетных мышц и тельцам Гольджи участвуют в системе обратных связей от двигательного аппарата.