

Иммунитет (определение). Врожденный и приобретенный (адаптивный) иммунитет. Структура иммунной системы.
Иммунитет(лат. immunitas — освобождение, избавление от чего-либо) — невосприимчивость, сопротивляемость организмаинфекциямиинвазиямчужеродных организмов (в том числе — болезнетворныхмикроорганизмов), а также воздействию чужеродных веществ, обладающихантигеннымисвойствами. Иммунные реакции возникают и на собственные клетки организма, измененные в антигенном отношении.
Обеспечивает гомеостаз организма на клеточном и молекулярном уровне организации. Реализуется системой иммунитета.
Биологический смысл иммунитета — обеспечение генетической целостности организма на протяжении его индивидуальной жизни. Развитие иммунной системы обусловило возможность существования сложно организованных многоклеточных организмов.
Иммунитет делится на врождённый и приобретенный.
Врождённый(неспецифический, конституционный) иммунитет обусловлен анатомическими, физиологическими, клеточными или молекулярными особенностями, закрепленными наследственно. Как правило, не имеет строгой специфичности к антигенам, и не обладает памятью о первичном контакте с чужеродным агентом. Например:
Все люди невосприимчивы к чуме собак.
Некоторые люди невосприимчивы к туберкулёзу.
Показано, что некоторые люди невосприимчивы к ВИЧ.
Приобретенныйиммунитет делится на активный и пассивный.
Приобретенный активныйиммунитет возникает после перенесенного заболевания или после введениявакцины.
Приобретенный пассивныйиммунитет развивается при введении в организм готовых антител в видесывороткиили передаче их новорожденному смолозивомматери или внутриутробным способом.
Также иммунитет делится на естественный и искусственный.
Естественныйиммунитет включает врожденный иммунитет и приобретенный активный (после перенесенного заболевания). А также пассивный при передаче антител ребёнку от матери.
Искусственный иммунитетвключает приобретенный активный после прививки (введение вакцины) и приобретенный пассивный (введение сыворотки)
Выделяют центральные и периферические органы иммунной системы. К центральным органам относят красный костный мозгитимус, а к периферическим -селезенку,лимфатические узлы, а также местноассоциированную лимфоидную ткань: бронхассоциированную (БАЛТ), кожноассоциированную (КАЛТ), кишечноассоциированную (КиЛТ,пейеровы бляшки).
Красный костный мозг— центральный орган кроветворения и иммуногенеза. Содержит самоподдерживающуюся популяциюстволовых клеток. Красный костный мозг находится в ячейках губчатого вещества плоских костей и в эпифизах трубчатых костей. Здесь происходит дифференцировкаВ-лимфоцитовиз предшественников. Содержит такжеТ-лимфоциты.
Тимус— центральный орган иммунной системы. В нем происходит дифференцировка Т-лимфоцитов из предшественников, поступающих из красного костного мозга.
Лимфатические узлы— периферические органы иммунной системы. Они располагаются по ходу лимфатических сосудов. В каждом узле выделяют корковое и мозговое вещество. В корковом веществе есть В-зависимые зоны и Т-зависимые зоны. В мозговом есть только Т-зависимые зоны.
Селезёнка— паренхиматозный зональный орган. Является самым крупным органом иммунной системы, кроме того, выполняет депонирующую функцию по отношению к крови. Селезёнка покрыта капсулой из плотной соединительной ткани, которая содержит гладко-мышечные клетки, позволяющие ей при необходимости сокращаться. Паренхима представлена двумя функционально различными зонами: белой и красной пульпой.Белая пульпасоставляет 20 %. Представлена лимфоидной тканью. Здесь имеются В-зависимые и Т-зависимые зоны. И также здесь есть макрофаги.Красная пульпасоставляет 80 %. Она выполняет следующие функции:
Депонирование зрелых форменных элементов крови.
Контроль состояния и разрушения старых и повреждённых эритроцитов и тромбоцитов.
Фагоцитоз инородных частиц.
Обеспечение дозревания лимфоидных клеток и превращение моноцитов в макрофаги.
Функции иммунной системы, концепция иммунного надзора. Современные теории иммунитета. Клонально-селекционная теория Бернета, теория «сетей» Йерне.
Клонально-селекционная теория иммунитета Бернета.
Клонально-селекционная теорияимеет в виду то, что каждыйлимфоцитсинтезирует антитела только одной определенной специфичности. И эти антитела располагаются на поверхности этого лимфоцита в качестве рецепторов.
Как показывают опыты, все поверхностные иммуноглобулины клетки имеют одинаковый идиотип: когда растворимый антиген, похожий на полимеризованныйфлагеллин, связывается со специфической клеткой, то все иммуноглобулины клеточной поверхности связываются с данным антигеном и они имеют одинаковую специфичность то есть одинаковый идиотип.
Антиген связывается с рецепторами, затем избирательно активирует клетку с образованием большого количества антител. И так как клеткасинтезирует антитела только одной специфичности, то этаспецифичностьдолжна совпадать со специфичностью начального поверхностного рецептора.
Специфичность взаимодействия антител с антигенами не абсолютна, они могут в разной степени перекрестно реагировать с другими антигенами. Антисыворотка, полученная к одному антигену, может реагировать с родственным антигеном, несущим одну или несколько одинаковых или похожихдетерминант. Поэтому каждое антитело может реагировать не только с антигеном, который вызвал его образование, но и с другими, иногда совершенно неродственными молекулами. Специфичность антител определяется аминокислотной последовательностью их вариабельных областей.
Клонально-селекционная теория:
Антитела и лимфоциты с нужной специфичностью уже существуют в организме до первого контакта с антигеном.
Лимфоциты, которые участвуют в иммунном ответе, имеют антигенспецифические рецепторы на поверхности своей мембраны. У B-лимфоцитоврецепторы- молекулы той же специфичности, что и антитела, которые лимфоциты впоследствии продуцируют и секретируют.
Любой лимфоцит несет на своей поверхности рецепторы только одной специфичности.
Лимфоциты, имеющие антиген, проходят стадиюпролиферациии формируют большой клон плазматических клеток .Плазматические клеткисинтезируют антитела только той специфичности, на которую был запрограммирован лимфоцит-предшественник. Сигналами к пролиферации служатцитокины, которые выделяются другими клетками. Лимфоциты могут сами выделять цитокины.
Первую, чисто биологическую, селективную теорию образования антител сформулировал Ерне, он предположил, что в организме синтезируется полный набор антител, но каждое из них образуется в небольшом количестве и независимо от какого-либо стимула поступает в кровь в виде естественных антител. Функция их состоит в том, чтобы избирательно связываться с соответствующим антигеном и таким способом доставлять его неким клеткам организма, для которых они служат сигналом к воспроизведению таких же молекул, т.е. к образованию антител. С этой точки зрения вторичный антительный ответ получал простое объяснение как результат появления после первой иммунизации увеличенного числа антител-носителей, присутствие которых будет одновременно способствовать селекции антигеном антител с более высоким сродством, что объясняет изменение качества антител при многократной иммунизации. Это была первая теория, которая объясняла также феномен иммунологической толерантности, принимая, что любые естественные антитела направленные против собственных антигенов будут немедленно абсорбироваться тканями организма, и таким образом не могут запустить образование аутоантител. Теория естественного отбора, предложенная Эрне, привлекла на свою сторону лишь немногих приверженцев инструктивных теорий, однако она имела большое историческое значение, т.к. дала стимул для теоретиков биологического направления таких как Вернет. Он использовал представления Эрлиха и Эрне о предсуществовании антител разной специфичности, но указывал на то, что каждое специфическое антитело синтезируется отдельным клоном клеток. По Бернету, при дифференцировке лимфоцитов от стволовой кроветворной клетки и при параллельном процессе мутационных изменений в генах, контролирующих синтез специфических антител возникают клоны клеток, которые способны взаимодействовать только с антигеном соответствующей специфичности. В результате такого взаимодействия формируется отобранный по специфичности клон, который либо секретирует антитела заданной специфичности, либо обеспечивает строго специфическую клеточную реакцию. Клональный селекционный признак организации иммунной системы выдвинутый Бернетом полностью подтвердился в настоящее время. Недостатком теории является представление о том, что многообразие антител возникает только за счет мутационного процесса.
Основным вкладом Й. в иммунологию стала теория «сетей», представленная им в 1974г. На сегодняшний день это самая детально разработанная и логичная концепция, объясняющая процессы мобилизации организма на борьбу с болезнью, а затем, когда болезнь побеждена, его возвращение в неактивное состояние. Е. подчеркивал, что разновидностей антител гораздо больше, чем белков, и «в динамическом состоянии наша иммунная система в основном замкнута на самой себе, вырабатывая антиидеотипные антитела к собственным антителам (т.е. антитела, соответствующие антигенному профилю собственных антител)». Поэтому реакция на инородное тело – это не просто усиленное образование того или иного антитела, а определенное «нарушение порядка» в чрезвычайно сложной саморегулирующейся системе.
Мукозоассоциирования лимфоидная ткань, её строение, функции. Секреторный IgA, строение, функции.
Ткань лимфоидная мукозассоциированная(МАЛТ) — лимфоидные ткани слизистых желудочно-кишечного, дыхательного и урогенитального трактов. Под слоем мукозного эпителия располагаются Пейеровы бляшки и лимфатические узлы.
В тропических странах тоже есть экономика и там можно вести дела. Открыть бизнес на Пхукетеможет любой предприимчивый человек.
Эти лимфоидные образования и объединяются под общим названием лимфоидная ткань, ассоциированная со слизистыми. В слизистой кишечника содержатся специализированные М-клетки, обеспечивающие транспорт антигенов от базальной мембраны в субэпителиальную область.
В последней локализуется большое количество лимфоцитов, особенно гамма/дельта CD8+ Т-клеток. В ламина проприа имеются все типы Т- и В-лимфоцитов, особенно IgA-синтезирующих, а также макрофагов и тучных клеток. В двух метрах слизистой кишечника содержится столько же лимфоцитов, сколько в совокупности в костном мозге, селезенке и лимфатических узлах. Иммунный ответ на инфекционные агенты, репродуцирующиеся в слизистых, развивается в основном по гуморальному типу.
В-лимфоциты дифференцируются и превращаются в плазматические клетки, продуцирующие специфические антитела, относящиеся к иммуноглобулинам разных классов. В случае образования иммуноглобулинов А они поступают в кровь или транспортируются через слизистую в форме секреторного sIgA, обеспечивающего противоинфекционную защиту слизистых. Избыточное образование антител IgE изотипа может привести к развитию аллергических реакций ГНТ I типа.
IgAсывороточный IgA составляет 15-20 % всей фракции иммуноглобулинов, при этом 80 % молекул IgA представлено в мономерной форме у человека. Секреторный IgA представлен в димерной форме в комплексесекреторным компонентом, содержится в серозно-слизистых секретах (например вслюне, слезах,молозиве,молоке, отделяемом слизистой оболочки мочеполовой и респираторной системы). Содержит 10-12 % углеводов, молекулярная масса 500 кДа.
![]()
|
Иммуноглобулины А (IgA)циркулируют в сыворотке крови (составляет 15-20% от всех Ig), а также секретируются на поверхность зпителиев. Присутствуют в слюне, слёзной жидкости, молоке и на поверхности слизистых оболочек.
М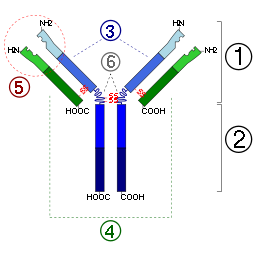 олекулыIgAучаствуют в реакциях
нейтрализации и агглютинации возбудителей.
Кроме того, после образования комплекса
Аг-АТ они участвуют в активации комплемента
по альтернативному пути.
олекулыIgAучаствуют в реакциях
нейтрализации и агглютинации возбудителей.
Кроме того, после образования комплекса
Аг-АТ они участвуют в активации комплемента
по альтернативному пути.
Дефицит IgA является одним из некорригируемых первичных дефектов иммунитета. Исправление данного дефекта состоит в лечении вторичных осложнений аллергической, аутоиммунной, либо инфекционной природы в совокупности с активацией сохранных элементов иммунитета с целью перекрытия недостатка продукции IgA (компенсации). Показаниями к проведению подобной иммуностимуляции являются преимущественно клинические проявления сниженной противоинфекционной сопротивляемости.
Известно, что уровень защиты от локальной вирусной инфекции желудочно-кишечного и респираторного трактов зависит не столько от наличия в организме сывороточного IgG к пневмотропным энтеропатогенным вирусам, сколько от содержания секреторного специфического иммуноглобулина A.
Отчетливый аффинитет к поверхностям слизистой оболочки, преобладание в секрете молочных желез, стабильная структура определяют биологическую значимость секреторного иммуноглобулина A в процессах защиты организма от агрессивного воздействия всевозможных патогенных агентов, в частности, вирусов.
Иммуноглобулин A в димерном виде синтезируют в lamina propria-клетках, а после связывания IgA с синтезированным в клетках эпителия иммуноглобулиновым рецептором транспортируют к поверхности слизистой оболочки. В момент появления IgA в просвете кишечника происходит частичное расщепление рецептора, вследствие чего IgA обогащается его фрагментом, именуемым секреторным компонентом. Очевидно, что секреторный IgA – совместный продукт клеток двух типов – эпителиальных и плазматических.
Способность секреторного иммуноглобулина А образовываться как в димерной, так и в тетрамерной формах значительно повышает его вирусонейтрализирующие свойства. Секреторный компонент защищает иммуноглобулин А от расщепления протеолитическим ферментом, что является его существенным преимуществом перед другими антителами. Секреторный IgA способен нейтрализовать вирус, как в просвете кишечника, так и во время транспортировки его внутрь клетки. Сначала димер иммуноглобулина А нейтрализует вирус в подслизистой кишечника, затем связывается с рецептором и транспортирует вирус в просвет кишечника.
Единая структура секреторного компонента и димерных иммуноглобулинов класса А, соединенная j-цепью является уникальным примером эволюционной адаптации иммуноглобулина на слизистых покровах, следствием чего является эффективное функционирование при постоянном воздействии антигенов разнообразной природы.
В различные внешние секреты, в том числе в молоко, молекулы иммуноглобулина A проникают из двух основных источников. IgA, выделяемый респираторным и пищеварительным трактами, секретами молочной и слезной желез, а также слюной, образуют плазматические клетки. Однако иммуноглобулин A, обнаруженный во внешних секретах может иметь происхождение и системного характера. Такой IgA продуцируется в клетках слизистой оболочки одного органа, поступает в кровь и переносится ею к слизистым покровам другого органа.
Проникнув в пейерову бляшку антиген активирует В- и Т-лимфоциты, используя лимфатический проток, те проникают сначала в мезентериальный лимфоузел, после чего в кровь, затем в селезенку и опять в кровь. Происходит их селективная локализация в лимфообразованиях слизистых оболочек, а также в секреторных железах экзокринной природы: слезных, молочных и слюнных. При этом локализация лимфоцитов происходит по большей части между клетками эпителия слизистой оболочки, что обеспечивает иммунный ответ клетки. Локализация В-лимфоцитов наблюдается преимущественно в lamina propria, где происходит их дифференциация в плазматические клетки, а также синтезируется иммуноглобулин A.
В-лимфоциты, их функции. Рецепторы зрелого В-лимфоцита. Популяции В-лимфоцитов.
B-лимфоци́ты(B-клетки, отbursa fabriciiптиц, где впервые были обнаружены) — функциональный типлимфоцитов, играющих важную роль в обеспечениигуморального иммунитета.
У эмбрионовчеловека и других млекопитающих B-лимфоциты образуются впечении костном мозге изстволовых клеток, а у взрослыхмлекопитающих— в красномкостном мозге.
При контакте с антигеном или стимуляции со стороны T-клетокнекоторые B-лимфоциты трансформируются вплазматические клетки, способные к продукцииантител. Другие активированные B-лимфоциты превращаются вB-клетки памяти.
Маркеры в-клеток
Характерной особенностью В-клеток является наличие поверхностных мембрано-связанных антител, относящихся к классам IgM и IgD. В комплексе с с другими поверхностными молекулами иммуноглобулины формируют антиген-распознающий рецептивный комплекс, ответственный за узнавание антигена. Также на поверхности В-лимфоцитов расположены антигены МНС класса II, важные в кооперации с Т-клетками, также на некоторых клонах В- лимфоцитов присутствует CD5 маркер, общий с Т-клетками. Рецепторы C3b компонента комплемента(Cr1, CD35) и C3d (Cr2,CD21) имеют определённую роль в активации В-клеток. Следует отметить, что маркеры CD19, CD20 и CD22используются для идентификации В-лимфоцитов. Также на поверхности В-лимфоцитов обнаруженыFc рецепторы.
Лимфоциты происходят от полипотентных стволовых клеток, дающих также начало всемклеткамкрови. Дифференцировка стволовых клеток крови по эритроидному, миелоидному либо лимфоидному пути зависит от микроокружения (в случае птиц дифференцировка стволовых клеток в В-лимфоциты происходит в фабрициевой сумке, у млекопитающих в костном мозге, где также происходит дифференцировка по миелоидному и эритроидному пути). Дифференцировка В-лимфоцитов условно делится на две стадии —антигеннезависимую(в которую происходит перестройкагеновиммуноглобулинови ихэкспрессия) и антигензависимую (при которой происходит активация,пролиферацияидифференцировкав плазматические клетки).
Пре-В-Клетки-предшественники не синтезируют тяжёлых и лёгких цепей, содержат зародышевые H и L гены, но содержат антигенный маркер, общий с зрелыми пре-В-клетками.
Ранние пре-В-клетки — D-J перестройки в Н генах.
Поздние пре-В-клетки — V-DJ перестройки в Н генах.
Большие пре-В-клетки Н-гены VDJ-перестроены; в цитоплазме имеются тяжёлые цепи класса μ.
Малые пре-В-клетки — V-J перестройки в L генах; в цитоплазме имеются тяжёлые цепи класса μ.
Малые незрелые В-клетки — L гены VJ-перестроены; синтезируют Н и L- цепи; на мембране расположены иммуноглобулины.
Зрелые В-клетки- начало синтеза IgD.
В-клетки поступают из костного мозга во вторичные лимфоидные органы (селезёнкуилимфатические узлы), где происходит их дальнейшее созревание, антиген-презентирование, пролиферация и дифференцировка в плазматические клетки и В-клетки памяти.
Экспрессия всеми В-клетками мембранных иммуноглобулиновпозволяет осуществляться клональному отбору под действием антигена. При созревании, антиген-стимулировании и пролиферации существенно меняется набор маркеров В-клеток. По мере созревания В-клетки переключаются от синтезаIgMиIgDна синтезIgG,IgA,IgE(при этом у клеток сохраняется способность синтезировать такжеIgMи IgD- вплоть до трёх классов одновременно). При переключении синтеза изотипов антигенная специфичностьантителсохраняется. Различают:
Собственно В-клетки (ещё называемые «наивными» В-лимфоцитами) - неактивированные В-лимфоциты, не контактировавшие с антигеном. Не содержат тельца Голла, вцитоплазмерассеяны монорибосомы. Полиспецифичны и имеют слабое сродство к многим антигенам.
В-клетки памяти - активированые В-лимфоциты, посредством кооперации с Т-клетками вновь перешедшие в стадию малых лимфоцитов. Являются долгоживущим клоном В-клеток, обеспечивая быстрый иммунный ответ и выработку большого количества иммуноглобулинов при повторном введении того же антигена. Названы клетками памяти, так как позволяют иммунной системе «помнить» антиген на протяжении многих лет после прекращения действия антигена. В-клетки памяти обеспечивают долговременный иммунитет.
Плазматические клетки - являются последним этапом дифференцировки активированных В-клеток, провзаимодействовавшими с антигеном. В отличие от остальных В-клеток несут мало мембранных антител и способны секретировать растворимые антитела. Являются большими клетками с эксцентрично расположенным ядром и развитым синтетическим аппаратом - шероховатый эндоплазматический ретикулумзанимает почти всю цитоплазму, также развит иаппарат Гольджи. Являются короткоживущими клетками (2-3 дня) и быстро элиминируются при отсутствии антигена, вызвавшего иммунный ответ.
Иммуноглобулины (антитела). Строение, классы и субклассы иммуноглобулинов
Антитела(иммуноглобулины, ИГ, Ig) — это особый классгликопротеинов, присутствующих на поверхностиВ-клетокв видемембраносвязанных рецепторови всыворотке кровии тканевой жидкости в виде растворимых молекул. Они являются важнейшим фактором специфическогогуморального иммунитета. Антитела используются иммунной системой для идентификации и нейтрализации чужеродных объектов — например, бактерий и вирусов. Антитела выполняют две функции:антиген-связывающую и эффекторную (вызывают тот или инойиммунный ответ, например, запускают классическую схему активациикомплемента).
Антитела синтезируются плазматическими клетками, которыми становятсяВ-лимфоцитыв ответ на присутствие антигенов. Для каждого антигена формируются соответствующие ему специализировавшиеся плазматические клетки, вырабатывающие специфичные для этого антигена антитела. Антитела распознают антигены, связываясь с определённымэпитопом— характерным фрагментом поверхности или линейной аминокислотной цепи антигена.
Антитела состоят из двух лёгких цепей и двух тяжелых цепей. У млекопитающих выделяют пять классов антител (иммуноглобулинов) — IgG, IgA, IgM, IgD, IgE, различающихся между собой по строению и аминокислотному составу тяжёлых цепей и по выполняемым эффекторным функциям.
Антитела являются относительно крупными(~150 кДа— IgG)гликопротеинами, имеющими сложное строение. Состоят из двух идентичныхтяжелых цепей(H-цепи, в свою очередь состоящие из VH, CH1, шарнира, CH2 и CH3 доменов) и из двух идентичныхлёгких цепей(L-цепей, состоящих из VL и CL доменов). К тяжелым цепям ковалентно присоединены олигосахариды. При помощи протеазыпапаинаантитела можно расщепить на дваFab(англ.fragment antigen binding — антиген-связывающий фрагмент) и одинFc(англ.fragment crystallizable — фрагмент, способный к кристаллизации). В зависимости от класса и исполняемых функций антитела могут существовать как вмономернойформе (IgG, IgD, IgE, сывороточный IgA) так и волигомернойформе (димер-секреторный IgA, пентамер — IgM). Всего различают пять типов тяжелых цепей (α-, γ-, δ-, ε-и μ- цепи) и два типа легких цепей (κ-цепь и λ-цепь).
антиинфекционныеилиантипаразитарныеантитела, вызывающие непосредственную гибель или нарушение жизнедеятельности возбудителя инфекции либо паразита
антитоксическиеантитела, не вызывающие гибели самого возбудителя или паразита, но обезвреживающие вырабатываемые имтоксины.
так называемые «антитела-свидетели заболевания», наличие которых в организме сигнализирует о знакомстве иммунной системы с данным возбудителем в прошлом или о текущем инфицировании этим возбудителем, но которые не играют существенной роли в борьбе организма с возбудителем (не обезвреживают ни самого возбудителя, ни его токсины, а связываются со второстепенными белками возбудителя).
аутоагрессивные антитела, илиаутологичныеантитела,аутоантитела— антитела, вызывающие разрушение или повреждение нормальных, здоровых тканей самогоорганизмахозяина и запускающие механизм развитияаутоиммунных заболеваний.
аллореактивныеантитела, илигомологичныеантитела,аллоантитела— антитела против антигенов тканей или клеток других организмов того же биологического вида. Аллоантитела играют важную роль в процессах отторжения аллотрансплантантов, например, при пересадкепочки,печени,костного мозга, и в реакциях на переливание несовместимой крови.
гетерологичныеантитела, илиизоантитела— антитела против антигенов тканей или клеток организмов других биологических видов. Изоантитела являются причиной невозможности осуществления ксенотрансплантации даже между эволюционно близкими видами (например, невозможна пересадка печени шимпанзе человеку) или видами, имеющими близкие иммунологические и антигенные характеристики (невозможна пересадка органов свиньи человеку).
антиидиотипическиеантитела — антитела против антител, вырабатываемых самим же организмом. Причём это антитела не «вообще» против молекулы данного антитела, а именно против рабочего, «распознающего» участка антитела, так называемого идиотипа. Антиидиотипические антитела играют важную роль в связывании и обезвреживании избытка антител, в иммунной регуляции выработки антител. Кроме того, антиидиотипическое «антитело против антитела» зеркально повторяет пространственную конфигурацию исходного антигена, против которого было выработано исходное антитело. И тем самым антиидиотипическое антитело служит для организма фактором иммунологической памяти, аналогом исходного антигена, который остаётся в организме и после уничтожения исходных антигенов. В свою очередь, против антиидиотипических антител могут вырабатыватьсяанти-антиидиотипическиеантитела и т. д.
Гуморальный иммунный ответ, его стадии. Отличия при ответе на Т-лимфоцит-зависимые и Т-лимфоцит-независимые антигены.