

14. Гистология - Histology / Эмбриология, тератология и основы репродукции человека _ В.Н. Запорожан, В.К. Напханюк, Е.Л. Холодкова. - О._ ОГМУ, 2000. - 378 с
.pdfSpinnbarkeit), межменструальные боли (Mittelschmertz) и ба-
зальная температура (небольшое снижение утренней температуры с последующим подъемом на 0,3–0,5 °С в середине цикла — характерный признак повышения уровня прогестерона). Наиболее надежным, однако, является гормональный метод, в частности, определение пика ЛГ в моче.
Желтое тело
После овуляции разорвавшийся фолликул начинает быстро превращаться в так называемое желтое тело, которое является временной эндокринной железой. Стенка фолликула спадается, остаточная полость заполняется кровью, которая затем организуется новообразованной соединительной тканью. Зернистый слой пролиферирует, а его клетки начинают накапливать желтый пигмент и становятся лютеиновыми клетками. Они синтезируют достаточно прогестерона, чтобы обеспечить имплантацию и дальнейшее развитие эмбриона. Сквозь теку врастают кровеносные сосуды и мелкие клетки, а клетки теки могут увеличиться в размере (паралютеиновые клетки). Они синтезируют эстроген. Если оплодотворение не произошло, желтое тело подвергается инволюции в течение 2-х недель. Менструальное желтое тело хорошо заметно на поверхности яичника.
Если оплодотворение произошло, желтое тело беременности увеличивается в размере и продолжает функционировать многие недели, после чего подвергается медленному обратному развитию, но остается различимым еще достаточно долго. Это зависит от адекватной гонадотропной (лютеотропной) поддержки гипофиза и эмбриона (человеческий хорионический гонадотропин — чХГ). Первые несколько недель после оплодотворения желтое тело обеспечивает выработку необходимого количества прогестерона для подготовки эндометрия к имплантации, затем его функции берет на себя плацента. В дегенерирующем желтом теле (как после овуляции, так и во время беременности) лютеиновые клетки замещаются фиброзной тканью, вследствии чего формируется беловатый рубец (белое тело).
Удлинение жизненного цикла функционирующего желтого тела необходимо для установления и поддержания успешной беременности.
19
Оплодотворение
Оплодотворение представляет собой процесс, который начинается с момента контакта сперматозоида со вторичным ооцитом, а заканчивается объединением материнского и отцовского хромосомного материала в метафазе первого митотического деления зиготы. Возможно, ооцит выделяет в фолликулярную жидкость химическое вещество, которое привлекает сперматозоид (хемотаксис). Зигота — это фактически эмбрион на стадии одной клетки.
Термин “концептус” используется для обозначения суммарных “продуктов” оплодотворения, включая внезародышевые органы и сам эмбрион, на любой стадии его развития.
Период между овуляцией и оплодотворением занимает не более одного дня, поэтому термин “постовуляторный возраст” используют в эмбриологии. Оплодотворение обычно происходит в ампуле (латеральный конец) маточной трубы. Оно состоит из нескольких фаз: а) контакт сперматозоида с прозрачной зоной ооцита, внедрение одного или нескольких сперматозоидов в прозрачную зону и в ооплазму, разбухание головки сперматозоида, окончание мейоза с образованием второго полярного тельца, б) формирование мужского (из головки сперматозоида) и женского пронуклеусов (в оотиде) и с) начало первого митотического деления или дробления зиготы. В норме реакции прозрачной зоны и корковой части ооцита блокируют проникновение более чем одного сперматозоида (полиспермию). Два пронуклеуса не сливаются, однако их ядерные оболочки расплавляются и формируют пузырьки. Две группы гомологичных хромосом затем начинают перемещаться и образуют веретено первого дробления. Имеющиеся в каждом пронуклеусе наборы из 23-х хромосом соединяются в 46 хромосом зиготы. Таким образом, диплоидный набор восстанавливается и формируется эмбриональный геном. Синтез РНК во время образования пронуклеусов сменяется синтезом ДНК после оплодотворения.
Критериями, позволяющими считать, что оплодотворение произошло, являются: наличие двух или более полярных телец в перивителлиновом пространстве, присутствие двух пронуклеусов в ооплазме, наличие остатков жгутика оплодотворяющего сперматозоида в ооплазме, выход кортикальных гранул из ооплазмы в перивителлиновое пространство. Первое полярное тельце может разделиться на два. Крайне редко, но впол-
20

не возможно оплодотворение любого из имеющихся полярных телец. Происхождение двуяйцевых близнецов можно объяснить оплодотворением: а) двух яйцеклеток, б) двуядерной яйцеклетки, с) полярного тельца.
Термин “супероплодотворение” используется в том случае, если во время одного полового акта может произойти оплодотворение двух или более яйцеклеток из одного овуляторного цикла. Термин суперфетация употребляют тогда, когда оплодотворение яйцеклетки произошло после формирования зародыша в предыдущем цикле, и он уже находится в полости матки.
Партеногенез
Партеногенез — процесс образования зародыша с возможным его развитием во взрослый организм из женской гаметы, без участия какой бы то ни было части мужской гаметы (Beatty, Kaufman). Такие эмбрионы могут быть гаплоидными или диплоидными. Партеногенез изучался, главным образом, на инбредных линиях мышей, у которых начиналось развитие после имплантации. Однако человеческие “эмбрионы”, найденные в атретических овариальных фолликулах, и считавшиеся партеногенетическими, оказались всего лишь похожими на дробящиеся клетки, поэтому утверждение о том, что ребенок может произойти партеногенетически, должно использоваться очень осторожно.
1.3. РАННИЙ ЭМБРИОНАЛЬНЫЙ ПЕРИОД
Первым 3-м неделям развития посвящены отдельные главы, потому, что именно в этом периоде закладываются органы и системы зародыша. Зародышевые листки образуются в течение первых двух недель, а на 3-й неделе формируются все основные органы. Должно пройти 3 недели, чтобы зигота диаметром 0,1 мм выросла до усложненного тела зародыша длиной около 3 мм.
На 1-й неделе зародыш представляет собой массу клеток — морулу, в которой затем появляется полость и морула превращается в бластоцисту.
21
Первая неделя
Почему дробящийся эмбрион продвигается вдоль маточной трубы еще окончательно не выяснено, однако сокращения трубы и движения ее ресничек играют важную роль в этом процессе. Митотические деления продолжаются таким образом, что число клеток возрастает, а общий объем протоплазмы остается прежним. Эмбрион входит в матку на стадии как минимум 8–12 клеток, а эндометрий находится в начале секреторной фазы, что соответствует лютеиновой фазе овариального цикла. Каждая клетка морулы в этот период считается тотипотентной, т. е. способной в случае изоляции сформировать полноценный эмбрион. Разделение этих клеток является причиной почти 1/3 случаев развития монозиготных близнецов.
Зародышевый геном
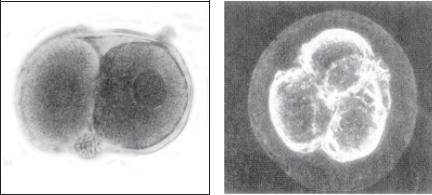
Начальное развитие зародыша происходит независимо от эмбрионального генома и находится под генетическим контролем материнского происхождения, который осуществляется посредством синтеза молекул РНК во время роста и созревания яйцеклеток. Уже на стадии 2-х клеток зародыш имеет ультраструктурные признаки (модификации клеточной мембраны) регионализации на поверхности бластомеров, по-видимому, под контролем информации, закодированной яйцеклеткой (возможно при помощи белков цитоскелета). Транскрипция генов
Рис. 1.3.1. Двухклеточный за- |
Рис. 1.3.2. Четырехклеточный |
родыш. Заметны полярные тельца |
зародыш |
и прозрачная оболочка |
|
22
человеческого эмбриона активируется вне ядра, как минимум, у 4-клеточного зародыша, но уровень синтеза РНК еще низкий. В 8-клеточном зародыше у некоторых бластомеров возрастает общая транскрипционная активность в результате действия как ядерных, так и внеядерных влияний. В этот период гены человеческого эмбриона и его генетическая уникальность отражаются в фенотипе (рис. 1.3.1, 1.3.2).
Приблизительно на стадии 16-клеточного зародыша, каждая клетка становится “детерминированной” либо к превращению в трофобласт, либо — в эмбриобласт (внутреннюю клеточную массу).
Приблизительно 4 дня
Полость бластоцисты (которая не является “бластоцелью”) появляется внутри клеточной массы на стадии приблизительно 32-х клеток. Четырехдневный эмбрион еще не прикрепился к слизистой оболочке матки.
Вскоре бластоциста высвобождается (“вылупливается”) из блестящей оболочки и в ней начинают дифференцироваться: 1) трофобласт (или трофэктодерма — расположенные по периферии зародыша клетки, образующие первый эпителий), и 2)
собственно эмбриональные клетки. Они, вначале в небольшом количестве, формируют внутреннюю клеточную массу, которая состоит из двух слоев: эпибласта и гипобласта. Сдвоенная пластинка эпибласта и гипобласта называется эмбриональным диском. Удвоение внутренней клеточной массы, возможно, является причиной образования монозиготных близнецов и, вероятно, это разделение происходит во время высвобождения из прозрачной зоны. Поверхность внутренней клеточной массы, прилежащая к трофобласту, определяет его дорзальную сторону, а та, что обращена к полости бластоцисты, обозначает вентральную. Если, как у мышей, гипобласт не принимает участия в образовании конечной зародышевой энтодермы, то на более поздних стадиях собственно зародыш полностью образуется из эпибласта внутренней клеточной массы. С вентральной стороны эпибласт лежит на базальной мембране (рис. 1.3.3).
Приблизительно 5–6 дней
Бластоциста начинает внедряться в стенку матки (эндометрий) и это является началом имплантации. Имплантация —
23
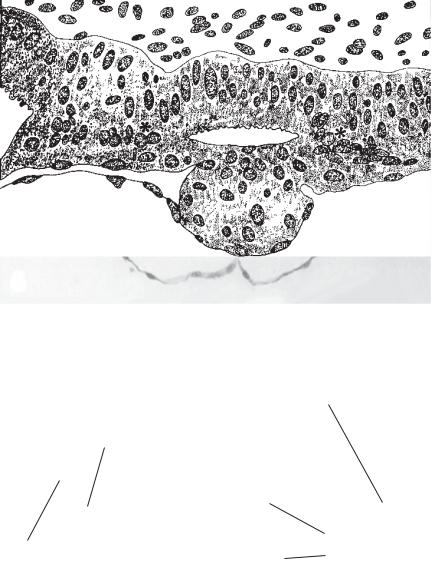
а
эндометрий
синцитио- |
маточный |
|
эпителий |
||
трофобласт |
||
эпибласт |
||
цитотрофобласт |
||
гипобласт |
||
|
б
Рис. 1.3.3. а — срез бластоцисты в начале имплантации. Видны две зоны адгезии между синцитиотрофобластом и эндометрием; б — двухслойный зародышевый диск и две зоны адгезии (отмечены звездочками)
24
процесс, в результате которого образуются специализированные тесные клеточные контакты между трофобластом и эндометрием (или другой тканью, в случае эктопической беременности). Имплантация включает в себя расплавление прозрачной зоны, адгезию бластоцисты к эндометрию, проникновение трофобласта и продвижение бластоцисты вглубь эндометрия. Позднее бластоциста оказывается полностью погруженной в оболочку эндометрия.
Возможно, имплантация происходит вследствие внедрения выростов трофобласта между интактными клетками эндометрия и, конечно, вследствие слияния клеток трофобласта и эндометрия. Имплантация — необычный процесс, ибо: 1) имеет сходство с инвазией злокачественных новообразований, 2) включает толерантность эндометрия (“хозяина”) к генетически чужеродным клеткам (трофобластическая “прививка”) и 3) является началом парадоксального слияния апикальных поверхностей клеточных мембран трофобласта и эндометрия
(Denker).
Стенка матки, в которую имплантируется эмбрион, состоит из трех оболочек: эндометрий (слизистая), миометрий (мышечная) и периметрий (серозная). Из них наибольшее значение в период имплантации имеет эндометрий, находящийся в этот момент в секреторной фазе, т. е. содержит активно функционирующие железы, большой запас питательных веществ, спиральные артерии. Если имплантация произошла, действие желтого тела (продуцирует прогестерон) сохраняет эндометрий; если имплантация не произошла, эндометрий разрушается в конце маточного цикла (менструация).
Успех имплантации также зависит от способности трофобласта вырабатывать иммуносупрессивные факторы, подавляющие материнский иммунный ответ. Она не произойдет в случае, если иммунная система матери отвергнет генетически чужеродный эмбрион.
Вторая неделя
На 2-й неделе имплантация завершается. В трофобласте, который вначале имел однородную плотную консистенцию, вскоре появляются лакуны и начинают образовываться ворсины хориона. Развиваются амниотическая полость и пупочный пузырек (желточный мешок). Некоторые дорзально расположенные клетки эмбрионального диска движутся в вент-
25
ральном направлении (кзади от эмбриона), формируя первичную полоску, которая определяет билатеральную симметрию зародыша. На 8-й день начинает определяться чХГ. Имплантация является триггером для начала синтеза чХГ концептусом и гормон вначале выявляют в крови, а через несколько дней — в моче матери. На этом основан способ ранней диагностики беременности.
С прогрессированием имплантации становятся необходимыми факторы роста, часть из которых вырабатывается самим эмбрионом. Некоторые пептиды (инсулиноподобные факторы роста) имеют большое значение для митоза и дифференцировки во время первого триместра беременности.
Приблизительно 7–12 дней
Соединительнотканные клетки стромы эндометрия увеличиваются и заполняются гликогеном. Этот процесс называют де-
цидуальной реакцией, а эндометрий становится децидуальной оболочкой (от лат. — decidua — отпадающая; отпадает после рождения).
В результате пролиферации клеток экстраэмбриональной мезодермы в каудальной области образуется мезенхима. Трофобласт и внезародышевая мезодерма формируют хорион. Это
— одна из зародышевых оболочек, окружающих эмбрион. Хорион образует хорионическую полость, в которой располагается зародышевый диск (рис. 1.3.4, 1.3.5).
К концу 1-й недели эмбрионы могут находиться на различных этапах имплантации: присоединения, внедрения и погружения бластоцисты в толщу эндометрия. Это занимает приблизительно 1 неделю. Трофобласт дифференцируется в двух главных направлениях: образование цитотрофобласта ближе к зародышевому диску и синцитиотрофобласта, являющегося производным цитотрофобласта и расположенного ближе к периферии. Трофобласт вначале плотный, однородный, затем в нем начинают образовываться заполненные кровью полости — лакуны, которые вскоре соединяются с сосудами эндометрия. На срезе маточные железы имеют вид “пилы”. Участки цитотрофобласта вдаются в синцитиотрофобласт и достигают центра мезодермы.
Амниотическая полость появляется, вероятно, во внутренней клеточной массе, возможно, в результате перераспределения клеток эпибласта. Она связана с эпибластом и дорзально
26

амнион
эпибласт
АП
энтодерма
желточный мешок
а |
б |
Рис. 1.3.4. а — на поперечном срезе двухнедельного зародыша видны амниотическая полость, зародышевый диск и желточный мешок. б— срез, проведенный более каудально. Можно различить первичную полоску и соединительный стебелек (на вершине). АП — амниотическая полость
дорзальная |
внезародыше- |
|
вая мезодерма |
||
|
||
|
зародыше- |
|
дорзальная |
вый диск |
|
|
||
вентральная пупочный |
|
|
стебелек |
|
|
левая |
|
|
ростральная |
|
|
дорзальная |
|
каудальная
а
правая
боковая
б
Рис. 1.3.5. а — зародышевый диск в 4 дня: имеются дорзальная и вентральная поверхности; б — отсутствие двусторонней симметрии к 13 дню
27
— со слоем амниотической эктодермы. (Амнион, как и хорион, является одной из зародышевых оболочек). На вентральной стороне зародышевого диска экстра-эмбриональная энтодерма растет, окружая полость, которая называется первичным пупочным пузырьком (желточным мешком). Зародышевый диск все еще остается двухслойным — состоит из эпибласта и энтодермы, являющейся производным гипобласта.
Монозиготные близнецы могут образовываться, начиная со стадии 2-х клеток и до стадии раннего зародышевого диска, пока еще не определилась билатеральная симметрия эмбриона.
Приблизительно 13 дней
Децидуальная оболочка дифференцируется на три зоны: 1) decidua basalis, расположенная в самой глубокой части эндометрия под местом имплантации; 2) decidua capsularis, лежащая между имплантировавшимся плодным яйцом и полостью матки на остатке хорионического мешка; 3) decidua parietalis
— оставшийся эндометрий, выстилающий полость матки, исключая зону имплантации.
Мезодермальные выросты, покрытые цитотрофобластом, формируют первичные ворсины хориона. Изначально их дистальные концы не являются свободными.
Амниотическая эктодерма вскоре достигает наружного слоя мезодермы. Вторичный пупочный пузырек развивается из первичного, вероятно, при спадении последнего и последующего вылущивания дистальной части. Каудальный конец зародыша связан с хорионом при помощи пупочного стебелька (будущего пупочного канатика).
Зародышевый диск слегка вытянут. Эпибласт представляет собой псевдомногослойный призматический эпителий. Определенные каудально расположенные клетки эпибласта мигрируют вентрально вдоль медиальной оси и формируют то, что называется первичной полоской, которая, как считают, является результатом взаимодействия эмбриональной энтодермы и плюрипотентных клеток эпибласта. Оставшиеся клетки эпибласта на дорзальной стороне зародыша составляют эмбриональную эктодерму.
Первичная полоска является местом, где клетки вдавливаются, пролиферируют и мигрируют, формируя эмбриональную мезодерму и энтодерму. Ростральный конец первичной полоски выделяется, образуя первичный узелок, который содержит
28