

14. Гистология - Histology / Эмбриология, тератология и основы репродукции человека _ В.Н. Запорожан, В.К. Напханюк, Е.Л. Холодкова. - О._ ОГМУ, 2000. - 378 с
.pdfсистема, например, головной и спинной мозг, окружена оболоч-
кой, названной менингеальной.
Гистогенез в нервной трубке
Призматическая эктодерма нервной трубки пролиферирует
иформирует псевдомногослойный эпителий, в котором ядра расположены на разных уровнях. Клетки, названные герминативными или желудочковыми, дают начало нейронам и глиальным клеткам и формируют герминативный матрикс или желудочковую зону. Следует отметить, что глиальные клетки не являются производными гистологически определенных элементов, формально называемых “спонгиобластами”. Нейроны и глиальные клетки происходят из одного и того же эпителия. Недавно было установлено, что, например, у крыс нейроны и клетки глии имеют общую клетку-предшественницу в желудочковой зоне. Желудочковые клетки выселяются из крайней пограничной сети, расположенной внутри (за полостью нервной трубки), и достигают базальной мембраны. Ядра изменяют свои позиции в процессе митотического цикла. Они мигрируют внутрь, где происходит митоз. После митоза ядра дочерних клеток отодвигаются от полости во время интерфазы и возвращаются обратно во время профазы. Чуть позже некоторые нейроэктодермальные клетки отделяются и свободно мигрируют.
Вначале внутренняя стенка нервной трубки состоит из желудочкового слоя. Затем свободная от ядер краевая зона перемещается кнаружи от желудочковой зоны и развивается промежуточная зона, соответствуя приблизительно тому, что называют мантийным слоем. Последняя содержит ядра постмитотических клеток — незрелых нейронов. Они не способны
кмитозу (поэтому термин “нейробласты” некорректен). Глиальные клетки, напротив, продолжают делиться и после того, как они мигрировали с желудочковой поверхности. Позже, в позднем зародышевом периоде, а также в плодном, пролиферация нейронов происходит не только в желудочковом слое, но
ив более периферически расположенных участках, называемых
вторичными пролиферативными зонами.
Существующая концепция “изнутри—наружу” отражает топографию нейронов, берущих начало от желудочковых клеток. В соответствии с этим правилом самые старые нейроны располагаются возле полости желудочка, а более молодые —
79
на периферии стенки мозга. Исключением из этой схемы являются самые зрелые клетки будущих полушарий мозга, которые обнаружены на периферии, кнаружи от кортикальной пластинки, а также в центре.
Нейроны
По мере формирования нейронов происходит значительное развитие гранулярной эндоплазматической сети. В цитоплазме появляются гранулы хроматофильной субстанции (Ниссля), становятся различимыми нейротрубочки и нейрофиламенты. Большинство нейронов формируется пренатально, тогда как образование глии в значительной мере происходит и в постнатальном периоде. В плодном периоде можно различить два больших периода образования нейронов: первый — с 15-й до 20-й недели, а второй — приблизительно в 25 недель.
Запрограммированная смерть незрелых нейронов во время развития является важным морфогенетическим явлением. При этом число молодых нейронов уменьшается, а количество пресинаптических нейронов соответствует количеству постсинаптических. В целом, число нейронов регулируется по принципу периферической обратной связи. После рождения дважды происходит снижение числа нейронов: 1) в 6 месяцев и вновь между 1 и 2 годами, это вероятно, связано с ростом коры мозга в этом периоде (количество нейронов остается неизмененным); 2) заметное и значительное снижение количества нейронов начинается после 2-х лет и длится до 16.
Нервный импульс по аксону движется от тела нейрона, а по дендриту — к телу. Клетки, дающие начало нейронам, вначале не имеют отростков, а затем становятся униполярными, когда начинает формироваться аксон, биполярными, когда возникает зачаток дендрита, и, наконец, мультиполярными (с одним аксоном и несколькими дендритами). Нейроны можно классифицировать:
1.Униполярные нейроны имеют лишь один аксон. Они достаточно многочисленны во время развития и крайне редко наблюдаются после рождения ребенка. Это палочковые и колбочковые клетки сетчатки.
2.Биполярные нейроны остаются биполярными. Они находятся в вестибулярном и улитковом (спиральном) узлах, обонятельном эпителии и сетчатке. Эти клетки имеют периферические отростки (дендриты) и центральный отросток (аксон).
80
3.Псевдоуниполярные нейроны. Во время развития два от-
ростка биполярного нейрона могут сходиться и, сливаясь, образовывать одинарный, короткий ствол, который дает начало миелиновым волокнам. Такие клетки находятся в спинномозговых и черепномозговых узлах.
4.Мультиполярные нейроны, большинство из которых образуют множество дендритов. Типичными примерами являются автономные узлы, спинной мозг (двигательные нейроны), кора головного мозга и мозжечка.
Нейроглия
Нейроглия или глия — интерстициальные клетки нервной системы не нервного происхождения. Глиальные клетки гораздо более многочисленны, чем нейроциты. Основными их типами являются: астроглия, олигодендроглия, микроглия, эпендима, звездчатые и клетки неврилеммы. Они начинают развиваться в желудочковом слое и вначале находятся в тесном контакте с желудочковой выстилкой. Это ранние радиальные глиальные клетки, играющие важную роль в определении направления миграции нейронов через стенку мозга.
Астроциты и олигодендроциты образуются из радиальных нейроглиальных клеток. Астроциты являются компонентами нейрогематического барьера. Нейроглия включает клетки неврилеммы периферических нервов, звездчатые клетки периферических ганглиев (те и другие происходят из нервного гребня) и эпендимоциты (выстилают центральный канал спинного мозга и полости желудочков мозга). Считается, что микроглия имеет мезодермальное происхождение. Нейролеммоциты (клетки Шванна) периферических нервов окружают часть одного аксона (в безмиелиновых нервах) или целый аксон, формируя несколько слоев (в миелиновых нервах). Звездчатые клетки (астроциты) периферической нервной системы окружают периферические клетки ганглиев.
Дифференцировка глиальных клеток происходит очень медленно, поэтому тяжело распознать, когда они впервые появляются. Однако при помощи иммунологических методов выделить глиальные клетки среди нервных можно в относительно раннем периоде. Первые клетки микроглии появляются в полушариях головного мозга в 5 недель. Образование глии длится всю жизнь. Большинство новообразований мозга имеют глиальное происхождение (глиома), чаще — астроцитарное (астроцитома).
81
Некоторые участки стенки нервной трубки остаются тонкими и выстланы эпендимоцитами — производными желудочковых клеток. Вдобавок специализированные эпендимоциты — клетки паутинного сплетения — вместе с богато кровоснабженной мягкой мозговой оболочкой формируют паутинные сплетения в желудочках мозга.
Можно предположить, что нейролеммоциты периферической нервной системы образуются как из нервной трубки, так и непосредственно из нервного гребня, который дает начало нейроцитам и глиальным клеткам автономной нервной системы и части узлов черепномозговых нервов.
Нервные волокна
Нервные волокна состоят из аксонов, окруженных оболочкой. В головном мозге человека волокна становятся видимыми в 3,5–4 недели, когда начинают формироваться первые проводящие пути. Примерно в это же время появляются периферические нервы: первыми — волокна подъязычного, затем вентральные и дорсальные корешки спинномозговых нервов.
По мере роста аксона из развивающегося нейрона его верхушка, имеющая коническую форму, совершает амебоидные движения, образуя конус роста. Принято считать, что растущие аксоны прокладывают себе путь к определенным участкам мозга вследствие воздействия химических веществ, продукция которых генетически детерминирована. Конус и короткие отростки (филоподии), захватывающие белки из внеклеточного пространства, содержат сеть микрофиламентов. Аксон растет в длину в определенном направлении и по пути формирует синаптические связи с определенными участками (чувствительными органами или мышцами). Прежде, чем установить связь с органом-мишенью, аксон должен расположиться возле нее. Прокладывание пути — сложный процесс, который зависит от сигналов с клеточных поверхностей, факторов диффузии и прорастания интерстиция вдоль аксона (под влиянием нейротропных факторов). Факторы окружающей среды, поведение, нервная (синаптическая) активность реr sе могут как пренатально, так и постнатально участвовать в формировании нервных связей взрослого человека.
В процессе иннервации мышц двигательными нейронами главная роль в поиске цели и формировании синапсов принадлежит конусу роста. Отрастание дендритов происходит позже,
82
после образования аксона. По мере роста дендрит образует конус роста, аналогичный тому, что образует аксон. Происходит ответвление большого количества дендритов, часть из которых затем рассасывается. Развитие дендритов связано с развитием синапсов. Образование отростков нейрона стимулируется определенными веществами, при этом основным яв-
ляется фактор роста нервов (ФРН).
Термин “синапс” обычно используют для обозначения соединения окончаний аксона (бутонов) с дендритами другого нейрона. Синапс содержит пресинаптические и постсинаптические элементы, а взаимодействия между ними устанавливаются в процессе развития. В двигательной зоне спинного мозга синапсы (преимущественно аксодендритические) определяются в 4,5 недели. В полушариях мозга синапсы определяют в 7 недель. В головном мозге млекопитающих происходит химическая передача импульса. Пресинаптическая луковица выделяет нейротрансмиттер, который проходит через синаптическую щель
исоединяется с молекулой рецептора на клеточной мембране постсинаптической клетки.
Цитоплазматические выросты нейролеммоцитов (в ПНС) или олигодендроцитов (в ЦНС) окружают аксон. Там, где происходит простое погружение, аксон называется безмиелиновым. Миелин представляет собой компактный слой, образованный многократно закручивающимся вокруг аксона выростом нейролеммоцита — мезаксоном. Процесс формирования миелина называется миелинизацией. При этом цитоплазма уплощается
иистончается так, что миелиновый слой состоит, главным образом, из клеточных мембран. Эти мембраны содержат билипидный слой, расположенный между двумя монослоями белка.
Миелинизация в ЦНС начинается сразу после того, как этот процесс начинается в ПНС, во втором триместре (вначале в задних и передних корешках спинномозговых нервов, затем в различных проводящих путях). Этот процесс длится несколько лет после рождения в постепенно замедляющемся темпе. Крупные чувствительные пути миелинизируются рано, в то время как к рождению ребенка полушария мозга содержат относительно небольшое количество миелина. Миелинизация становится особенно уязвимой ближе к рождению. Если установлено время и механизмы миелинизации, можно определить локализацию разных проводящих путей нервной системы. Множество восходящих и нисходящих нервных волокон распола-
83
гается в краевой зоне, образующей белое вещество. Тем не менее, проведение импульсов и функциональная активность проявляются еще до формирования миелиновой оболочки.
Периферические аксоны
Все периферические аксоны покрыты нейролеммоцитами, имеющими эктодермальное происхождение (нервный гребень). Во время развития они объединяют вырастающие аксоны и образуют миелиновую оболочку.
Вбезмиелиновых периферических нервах несколько аксонов вдавливаются в цитоплазму одного нейролеммоцита, однако при этом компактный слой миелина не образуется. Примерами могут быть постганглионарные автономные нервные волокна и некоторые волокна болевой чувствительности.
Вмиелиновом периферическом нерве каждый нейролеммоцит покрывает сегмент аксона. Область соединения двух сег-
ментов называется межузловым перехватом Ранвье. Базаль-
ная мембрана окружает нейролеммоцит и вместе с клеточной мембраной образует “неврилемму”, заметную при световой микроскопии.
Центральные аксоны
В ЦНС безмиелиновые аксоны проходят через массу (нейропиль) нейроглиальных клеток, дендритов и других аксонов. Вокруг некоторых аксонов формируется миелиновая оболочка, имеющая такое же строение, как и в ПНС. Один олигодендроцит может образовать миелиновую оболочку для нескольких аксонов. Преломляющие свойства миелина предопределяют цвет белого вещества головного мозга.
84

ГЛАВА 2
ЧАСТНАЯ ЭМБРИОЛОГИЯ
2.1. НЕРВНАЯ СИСТЕМА И ОРГАНЫ ЧУВСТВ
В 1920 г. Мангольд и Шпеман, исследовав зародыши амфибий, доказали, что нейральная детерминация является результатом взаимодействия предполагаемой нейральной эктодермы и будущей хордомезодермы (у млекопитающих соответствует ткани, развивающейся из первичной полоски). Эффект, когда формирование компетентной эктодермы индуцировано организатором к формированию нервной ткани, называется первич-
ной нейральной индукцией.
На наш взгляд, первичная индукция представляет собой процесс воздействия на эпибласт двух активных компонентов (повидимому, белковой природы), который приводит к началу образования нервной системы в ростро-каудальном направлении.
Фактор роста нервов, который можно обнаружить в подчелюстных слюнных железах, — пептид, стимулирующий формирование отростков (нейритов) от нейронов и (как нейротропин) поддерживает существование нейронов. Это необходимо для нормального развития нервной системы. Его рецептор является, вероятно, фотоонкогеном (называется trk). Факторы роста и нейротрансмиттеры генерируют сигналы, воздействуя на рост нервов и формирование нейритов; они также оказывают влияние на существование нейронов и формирование синапсов. Дисбаланс приводит к нейродегенерации, включающей такие специфические расстройства, как болезнь Альцгеймера. Эпидермальный фактор роста и рецепторы к нему возможно также имеют немаловажное значение в развитии нервной системы.
85
Формирование нервной трубки
Головной и спинной мозг, т. е. ЦНС, развиваются из участка эктодермы, названного нервной пластинкой, складки которой формируют нервные гребни. Из них в результате нейруляции формируется нервная трубка. На этой стадии развития зародыш иногда называют нейрулой.
Эксперименты на цыплятах показали, что нейруляция происходит пренодально, поэтому все отделы ЦНС развиваются рострально по отношению к первичному узелку. Первым видимым проявлением нервной системы у человека, тем не менее, является серединная — нервная бороздка, — которая начинает определяться у некоторых зародышей в 18 недель. Нервные складки, ограничивающие борозду с обеих сторон, вначале содержат больше материала будущего головного мозга, чем спинного. Вскоре появляется мезенцефальный перегиб. В результате можно различить три основных отдела головного мозга: передний (прозэнцефалон), средний (мезенцефалон) и задний (ромбэнцефалон). Нервная пластинка значительно удлиняется, коррелируя с длиной зародыша.
Первичная нейруляция
Прогибание нервной пластинки для формирования нервной трубки называется первичной нейруляцией. Этот процесс связан с характерными изменениями формы нейроэпителиальных клеток, когда они из кубических превращаются в клиновидные. Некоторые исследователи считают, что эти изменения вызваны движениями внутриклеточных микротрубочек и микрофиламентов. Более того, показано, например, на куриных эмбрионах, что изменения формы нейроэпителиальных клеток являются результатом индуцирующих взаимодействий с будущей нотохордой. Распространение внеклеточного матрикса мезенхимы может играть важную роль в углублении нервных складок. Факторы, необходимые для морфогенеза нервной системы, включают изменения формы клеток, их миграцию, дифференцированную пролиферацию и апоптоз.
В переднем мозге вскоре можно различить две составные части: маленькую — ростролатеральную (телэнцефалон) и большую — диэнцефалон, — преимущественную долю которого составляют оптическая и таламическая части. В оптической области начинает появляться оптическая бороздка, которая является первым признаком развивающегося глаза.
86
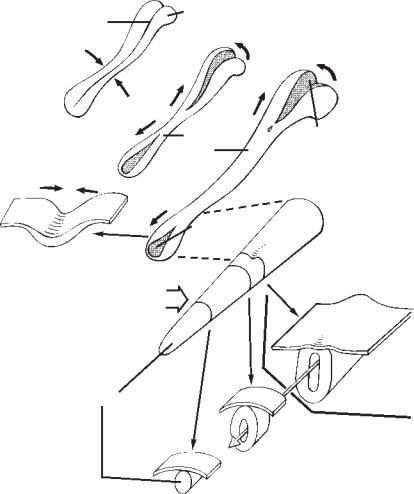
А
нервная бороздка
В′
каудальное
возвышение
нервный тяж
Ж
нервная
складка
Б
нервная
трубка
каудальный
нейропор
Е
В
ростральный
нейропор
Г
Д
первичная
нейруляция
вторичная
нейруляция
Рис. 2.1.1. Формирование нервной трубки. В 22 дня начинается слияние (Б) нервных складок (А). Вследствие слияния образуются два нейропора на ростральном и каудальном концах (В) зародыша, которые закрываются в 24 и 26 дней соответственно. На Г и Д небольшая ямка показывает место возникновения каудального нейропора, за пределами которого нервная трубка формируется путем вторичной нейруляции. Е — более рострально в тяже появляется полость, которая соединяется с той, что образовалась в результате первичной нейруляции
87
Замыкание нервной борозды начинается в месте соединения будущего головного и спинного мозга. Перед и после слияния поверхностных (эктодермальных) клеток обеих сторон, из места аналогичного слияния нейроэктодермальных клеток нервных складок выделяются клетки нервного гребня. Они развиваются из нейросоматического соединения (большей частью, по-видимому, непосредственно из нейральной эктодермы) и мигрируют в различные отделы организма.
Пока еще открытые концы развивающейся нервной труб-
ки называются ростральным и каудальным нейропорами. По-
лость трубки посредством нейропоров соединяется с амниотической полостью. Ростральный нейропор закрывается приблизительно на 24-й день, а каудальный — на 26-й. Закрытие рострального нейропора, в основном, идет в обоих направлениях — ростральном и каудальном. Окончательное закрытие рострального нейропора происходит, вероятно, на месте будущей комиссуральной пластинки, т. е. в середине зародышевой терминальной пластинки. Телэнцефалон все еще остается средней частью головного мозга: средний телэнцефалон. Характерной особенностью внутренней поверхности нервной трубки являются группы небольших вдавлений на каждой ее стороне, которые называются нейромерами. Спинномозговая часть нервной трубки быстро растет в длину.
Вторичная нейруляция
Каудальная часть нервной трубки развивается не в результате слияния нервных складок, а скорее из недифференциро-
ванной массы клеток — каудальной эминенции (или конечной почки). Как только каудальный нейропор закрывается, нервная ткань образует нейральный тяж, в который врастает полость уже сформированной нервной трубки, расположенной более рострально. Этот процесс называется вторичной нейруляцией. Он начинается в месте закрытия каудального нейропора, у которого уровень 31 сомита соответствует уровню будущего второго крестцового позвонка.
Оболочки мозга
В рыхлой соединительной ткани между зачатком скелета и нервной трубкой начинают появляться полости, уже различа-
88