

Micro-Nano Technology for Genomics and Proteomics BioMEMs - Ozkan
.pdfPEPTIDE ARRAYS IN PROTEOMICS AND DRUG DISCOVERY |
213 |
|||
library |
# of spots |
peptides/spot |
|
|
XXOOXX |
400 |
160,000 |
|
|
|
|
|
|
|
XODDOX |
400 |
400 |
|
|
|
|
|
|
|
ODDDDO |
400 |
1 |
|
|
|
|
|
|
|
DDDDDD |
|
|
|
|
FIGURE 7.44. Iterative deconvolution process of active peptide mixtures. A starting hexamer library of the type XXOOXX (X = randomized position; O = defined position) is screened and the best dipeptide combination OO is selected for the first deconvolution library (XODDOX; D = defined position identified from the preceding library). Subsequently, the second deconvolution library ODDDDO is based on the best tetrapeptide motif ODDO from the preceding library and leads to a single peptide (see 7.3.2.1.).
defined early in the process that have a more positive effect on those defined later in the deconvolution.
A dramatic increase in the effectiveness of peptide and peptide mixture multiple automated syntheses paved the way for more complex libraries of the type XXXXO1O2O3XXXX (8000 peptides mixtures) [496]. For example, the most complex library prepared by the SPOT technique described so far is of the type XXXX[3O3X]XXXX. The internal core [3O3X] is an abbreviation for three defined and three randomized positions arranged in all possible combinations XXXX[O1O2O3XXX]XXXX; XXXX[O1O2XO3XX]XXXX and so on [272]. This library comprised 68,000 spots and was used to identify not only antibody epitopes but also other peptides that bind to the antibody‘s paratope in a completely different way, referred to as mimotopes [177]. This was shown by structure determination of the peptide/antibody complexes by X- ray crystallography [255]. In many cases such complex libraries are essential for identifying peptide ligands that may require a certain number of key residues in a distinct pattern.
An alternative way to reduce the number of peptide mixtures that have to be prepared, yet match as many defined positions as practicable uses so-called combinatorial clustered amino acid peptide libraries [45, 271]. Each cluster contains physicochemically similar amino acids. The rationale of this approach is based on the assumption that physicochemically related amino acids contribute similarly to binding. For instance, grouping the amino acids into six clusters would lower the number of peptide mixtures in a combinatorial library containing four non-random positions from 204 (160,000, with four defined positions) to 64 (1,296, with four cluster positions). Kramer et al. described the epitope mapping of anti-transforming growth factor α (TGFα) mab Tab2 using a library of the type XC1C2C3C4X (C = one of six amino acid clusters [APG], [DE], [HKR], [NQST], [FYW], [ILVM]) in comparison to phage display techniques [271]. The peptide library array identified several motifs unrelated to the known TGFα-derived linear epitope sequence, whereas the phage display technique only revealed peptide ligands closely related to the wild-type epitope.
Several other combinatorial library techniques either as combinations or modifications of the principles described above or with unrelated design strategies were introduced for non-array technologies, but are similarly applicable for peptide arrays. A very interesting technique worth mentioning here is the so-called orthogonal library concept [111, 421]. The
214 |
ULRICH REINEKE, JENS SCHNEIDER-MERGENER AND MIKE SCHUTKOWSKI |
principle is that the same compound is represented in two different mixtures. Comparative activities of different mixtures observed after screening enables identification the compound responsible for activity.
In contrast to biological display techniques or in vitro translation systems, chemically prepared peptide arrays offer the opportunity to incorporate unnatural building blocks, e.g., D-amino acids, peptoidic building blocks, 1,3,5-trisubstituted hydantoins, 1,3,5-trisubstituted triazines, etc. A more detailed description of peptidomimetic classes applied so far is given in Section 7.5.5.
7.3.2.2. Random Libraries An alternative to protein sequence-derived or combinatorial peptide array libraries is to use sets of randomly generated peptide sequences. Recently, a peptide array approach was described using a library of 5520 randomly generated individual 15-mer peptides sequences prepared by SPOT synthesis and incorporating all genetically encoded amino acids except cysteine [458]. Of course, this only covers an extremely small fraction of the potential sequence repertoire. However, the peptide library array was successfully used to identify specifically binding peptide epitopes and mimotopes of three different antibodies (anti-IL-10 mab CB/RS/13, anti-TGFα mab Tab2, anti-p24 (HIV-1) mab CB4-1). Initially identified peptide ligands mostly had very low affinities for the antibodies with dissociation constants around 10−4 M. However, subsequent substitutional analyses revealed several analogs with dissociation constants in the low micromolar and high nanomolar range in a one step process. In two other studies 4450 randomly generated 12-mer peptides prepared on 10 “mini-Pepscan cards” (455 peptides per card) as well as a tripeptide library comprising the genetically encoded amino acids in all possible combinations were used to identify peptides binding to monoclonal antibodies against protein-S of transmissible gastroenteritis virus (TGEV) (mab 6A.A6 and 57.9), an EGF-like domain of the surface protein pfs25 of P. falciparum (mab 32F81), and the FLAG-tag (mab M2). Several peptides were identified as either homologous to the wild-type epitope sequence [516] or completely unrelated mimotopes [517]. Later, this approach was theoretically discussed and an algorithm to extract the amino acids required for binding was described [561].
In addition to randomly generated peptide library arrays, this approach was also used for peptidomimetics. Heine et al. described an 8000 membered hexapeptoid and hexapeptomer array used to identify peptidomimetics that bind the anti-TGFα mab Tab2 [202]. The best compound had a dissociation constant of 2.7 µM. The same antibody was used to probe an array of 8000 1,3,5-trisubstituted triazines, with the best hit having a dissociation constant of approximately 400 µM [488].
7.4. ASSAYS FOR PEPTIDE ARRAYS
This Section describes the general aspects of assay systems to probe peptide arrays. It is divided into the Sections “Screening” (4.1.) and “Read-out” (4.2.) addressing either the molecular recognition event, or how one observes which peptide was bound and/or converted by an interaction partner or enzyme. Of course, the quality of an assay system depends on the proper combination of screening and read-out methods.
PEPTIDE ARRAYS IN PROTEOMICS AND DRUG DISCOVERY |
215 |
A 

 B
B 
green fluorescent protein (GFP) fluorescence dye
radioactive label
chemoluminescent substrate
enzyme
 chromogenic substrate
chromogenic substrate
affinity purification tag or fusion moiety
C |
|
D |
|
|
E |
|
|
F |
|
|
G |
|
|
|
|
|
|
|
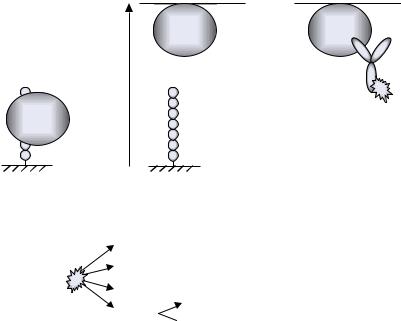
FIGURE 7.45. Binding assays and detection methods to identify peptide-protein interactions (see 7.4.1.1.).
(A) Antibody epitope mapping with a directly labeled antibody; (B) antibody epitope mapping using a labeled secondary antibody; (C) detection with a directly labeled protein; (D) immunological detection of peptide-protein interactions using a directly labeled primary antibody; (E) immunological detection of peptide-protein interactions using a primary antibody in combination with a labeled secondary antibody; (F) immunological detection of peptide-protein interactions using a labeled antibody directed against an affinity purification tag or a fusion moiety; and (G) immunological detection of peptide-protein interactions using an antibody directed against an affinity purification tag or a fusion moiety in combination with a labeled secondary antibody.
7.4.1. Screening
Molecular recognition events on peptide arrays include either ligand binding (see 7.4.1.1.) or enzymatic conversion (see 7.4.1.2.). Chemical transformations of peptides on arrays (see 7.5.4.5.) or binding of intact cells (see 7.5.4.6.) are only described briefly.
7.4.1.1. Ligand Binding As shown in Table 7.4, peptide arrays are most frequently applied to study peptide binding by polyclonal or monoclonal antibodies. This can be achieved with directly labeled [593] primary antibodies (Figure 7.45A) or with labeled secondary antibodies (Figure 7.45B). Proteins in general can also be labeled directly (Figure 7.45C). Alternatively, they are detected using an antibody against the protein itself (Figure 7.45D) or recognition of a purification tag (poly-His-tag, Strep-tag, etc.) or a fused region, e.g., an Fc or GST fusion (Figure 7.45F). Similar to detection of peptide-antibody binding, proteins and fusion proteins can be recognized with labeled primary as well as secondary antibodies (Figures 7.45E, G). Generally, labeled protein A or G can be used as an alternative to labeled secondary antibodies [578]. For all immunological detection systems it is critical to rule out direct, nonspecific binding of the detection antibodies (or protein A/G) to the peptides. Incubation procedures as well as control experiments for peptide arrays prepared by SPOT synthesis have been described and are analogous in principal for all types of arrays
216 |
ULRICH REINEKE, JENS SCHNEIDER-MERGENER AND MIKE SCHUTKOWSKI |
blotting membrane
electrotransfer
A B C
green fluorescent protein (GFP)
fluorescence dye
radioactive label
chemoluminescent substrate
enzyme
 chromogenic substrate
chromogenic substrate
FIGURE 7.46. Electrotransfer of peptide-bound proteins. (A) Protein binding to the peptide array; (B) Protein electrotransfer onto a nitrocellulose or polyvinylene difluoride membrane; and (C) detection after immobilization on the blotting membrane (see 7.4.1.1.).
[453]. In addition, false-positive results can occur when using fusion proteins, and this must be checked by control incubations with the fusion moiety alone.
In most cases both screening and read-out are carried out directly on the peptide arrays. Alternatively, peptide-bound antibodies or proteins can be electrotransferred onto nitrocellulose or polyvinylene diflouride (PVDF) membranes (Figure 7.46; [453, 481]). This procedure, which is only applicable for peptide arrays on porous membranes, results in an exact mirror image of the ligands captured on the array. Subsequently, incubation with detection antibodies for example, and read-out by any of the methods described below are carried out on the blotting membrane. This procedure was extensively applied for probing peptide arrays with chaperones (see 7.5.2.3.; [481]) and is especially well suited for mapping protein homodimerization sites [205, 266]. Here, a peptide scan of the chosen protein is incubated with the protein itself. Detection with an antibody against this protein would usually lead to false-positives due to detection antibody interactions with protein sequence-derived peptides on the array (i.e., antibody epitope mapping), a problem avoided on the electrotransfer membrane. In addition, the electrotransfer procedure is extremely useful for detecting low-affinity peptide-protein interactions. This is due to the binding equilibrium shifting towards un-complexed detection antibody or protein during subsequent array processing. The advantage of the electrotransfer procedure is that all incubation and washing steps are carried out after transfer and immobilization of the primary incubation molecule. Detailed experimental protocols are described elsewhere [453].
PEPTIDE ARRAYS IN PROTEOMICS AND DRUG DISCOVERY |
217 |
radioactive label |
|
Me |
Me |
 fluorescence dye
fluorescence dye
Me |
MeS |
Me |
A |
|
|
B |
|
|
C |
|
|
D |
|
|
E |
|
|
|
|
|
|
|
|
chelator
|
|
|
green fluorescent protein (GFP) |
|
|
|
fluorescence dye |
F |
G |
H |
radioactive label |
chemoluminescent substrate
enzyme
 chromogenic substrate
chromogenic substrate
FIGURE 7.47. Binding assays and detection of non-protein ligands binding to peptide arrays (see 7.4.1.1.).
(A) Binding of metal ions to peptide arrays is detected by radioactive isotopes; (B) detection via precipitating chromogenic chelators; (C) chelators that are labeled e.g., with peroxidase; (D) insoluble metal sulfides; (E) fluorescence dye labeled peptides. Other interactions identified on peptide arrays are (F) binding of labeled oligonucleotides;
(G) binding of dyeor peroxidase-labeled peptides; and (H) binding of heme to arrays of synthetic four helix bundle mini-proteins.
Non-protein ligands used to probe peptide arrays include metal ions, DNA, peptides and protein cofactors. Screening with metal ions is carried out with radioactive isotopes detected by autoradiography (Figure 7.47A), employing precipitating chelators or insoluble metal sulfides that are quantified by densitometry (Figure 7.47B and D), with chelators that are coupled to enzymes (peroxidase) or other markers for detection (Figure 7.47C), or with peptide coupled fluorescence labels like dansyl that show an increased fluorescence intensity upon metal ion (Pb2+, As3+) binding (Figure 7.47E) [172, 265, 269, 270, 346, 494]. Oligonucleotide binding to peptides is detected using 32P-labeled DNA or RNA (Figure 7.47F; [465, 466]). Peptide binding to protein domain arrays can be performed with peroxidase labeled [546, 547] or dye coupled peptides [548] (Figure 7.47G). Cofactor binding, for example heme binding to four-helix bundle protein arrays was carried out spectroscopically (Figure 7.47H; [441, 495]).
7.4.1.2. Enzymatic Conversion Measuring the activity of enzymes that modify peptides on arrays involves either enzymes adding something to the peptides (kinases, acetyl transferases, glycosyltransferases, etc.) or releasing a peptide part (proteases, phosphatases, etc.). In principle, all enzymes modifying peptides or proteins can be applied to screen peptide arrays. However, so far only assays for proteases, kinases, phosphatases, esterases and glycosyltransferases have been described, reflecting their respective importance in basic research and drug discovery. The different types of assays are described in detail below: kinases and phosphatases (7.5.3.1.; Figure 7.48) and proteases (7.5.3.2.; Figure 7.49). The binding specificity of proteins that convert or bind to peptides or proteins without covalent modifications, e.g., chaperones (see 7.5.2.3.) and isomerases (Table 7.4) is studied using binding assays as described above (see 7.4.1.1.).

218 ULRICH REINEKE, JENS SCHNEIDER-MERGENER AND MIKE SCHUTKOWSKI
A |
B |
kinase 32/33P |
kinase |
|
P |
[γ -32/33P] ATP |
ATP |
|
|
|
green fluorescent protein (GFP) |
|
fluorescence dye |
phosphate acceptor amino acid
radioactive label
chemoluminescent substrate
enzyme
 chromogenic substrate
chromogenic substrate
FIGURE 7.48. Kinase assay on peptide arrays (see 7.5.3.1.). (A) The array is incubated with the kinase of interest in the presence of [γ−32 or 33P]ATP and detection is performed by autoradiography. (B) Alternatively, the phosphorylation is measured with a labeled anti-phospho-amino acid antibody.
FIGURE 7.49. Different types of protease assays (see 7.5.3.2.). (A) Assay with N-terminally labeled immobilized peptides in the 96-well plate format [122]; (B) alternative assay with N-terminally labeled immobilized peptides in the 96-well plate format [254, 455]; (C) protease assay with internally quenched peptides on arrays [101, 254, 450]; (D) detection of peptide cleavage after electrotransfer of the released N-terminal peptide fragment [275];
(E) protease assay with peptides having a fluorogenic group C-terminal to the scissile bond [486, 616]; and (F) peptides with a fluorescence dye at the free terminus for array-based assays with decreasing signal upon cleavage.
PEPTIDE ARRAYS IN PROTEOMICS AND DRUG DISCOVERY |
219 |
7.4.2. Read-Out
Quantification of peptide-bound ligands or enzymatic peptide conversion employs chemoluminescent, fluorescent, radioactive, chromogenic, and label-free read-out methods such as surface plasmon resonance (SPR), mass spectrometry (MS), or atomic force microscopy (AFM). The following Sections describe these principles and discuss their advantages and drawbacks.
7.4.2.1.Chemoluminescence Detection of peptide-protein binding most often employs chemoluminescence read-out since a huge number of peroxidase-labeled monoclonal and polyclonal antibodies are commercially available. Very high sensitivity is achieved using a chemoluminescence substrate combined with either imaging systems or X-ray or photographic films. Luminol-based substrates can be mixed very cheaply or purchased from research reagent suppliers. In addition, ultra-high sensitivity substrates are commercially available. Fortunately, peroxidase itself shows almost no detectable binding to the peptides, and regeneration of peptide arrays, which is often done with SPOT synthesis-prepared arrays, is rather easy since no precipitates accumulate on the array support as occurs with chromogenic read-out methods (see 7.4.2.4.). Signal amplification and increased sensitivity is achieved by coupling more than one enzyme to the analyte or detection molecule, e.g., a second antibody or protein A or G.
7.4.2.2.Fluorescence The sensitivity of fluorescence read-out depends on the number of fluorescent moieties coupled per analyte molecule, the quantum yield of the fluorescent dye, the peptide loading, and the amplification achieved, for example by a sandwich assay with primary and secondary antibody. Fortunately, many secondary antibodies labeled with different fluorescence dyes are available. However, background fluorescence from the array support can be a severe drawback. Such background signals could be almost completely suppressed using planar waveguide technology in combination with fluorescencebased detection methods [400, 401, 584]. Glass chips usually have significantly lower intrinsic background compared to the cellulose or polypropylene membranes commonly used for arrays prepared by the SPOT technology. These porous membranes usually contain traces of fluorescent substances left over from the production process. Another source of background fluorescence can arise from the peptides, peptidomimetics or often side products from the synthesis process, e.g., side chain protecting groups. Generally, fluorescent
dyes with longer emission wavelengths, i.e., in the red range of the visible spectrum, such as Texas Red R are preferable to avoid interference with background fluorescence from substances in complex mixtures such as cell lysates. If the peptides on the array are labeled themselves, as required for some protease assays (see 7.5.3.2.; Figure 7.49), bulky and often hydrophobic fluorescence labels with longer emission wavelengths can cause severe problems by affecting the analyte’s interaction (e.g., a protease) with the peptides. In such cases aminobenzoic acid, with a molecular weight in the average range of natural amino acids, is often used in order to reduce assay and read-out artifacts [102, 254, 455].
7.4.2.3.Radioactivity Binding assays with radioactively labeled screening molecules are often used due to their very high detection sensitivity. Generally, proteins can be labeled with 125I [126, 582], 35S [44], 14C [377], or phosphorylated with [γ-32/33P]ATP
220 |
ULRICH REINEKE, JENS SCHNEIDER-MERGENER AND MIKE SCHUTKOWSKI |
[75, 139, 539]. The major advantage compared to immunological detection methods is that no controls are needed to rule out false-positive results from detection antibodies binding directly to the peptides. The drawback is that some proteins are denatured or modified in their activity or binding properties by 125I-labeling or 32P/33P phosphorylation, necessitating a control activity test with the radioactive protein.
Phosphorylation of peptides by kinases is the most important radioactivity-based assay (Figure 7.48A). Peptide arrays with a phosphate acceptor amino acid are incubated with the kinase of interest in the presence of [γ-32/33P]ATP (see 7.5.3.1.; [50, 143, 330, 338, 395, 484, 503, 539]). Incorporated radioactivity is measured using a phosphoimager or by exposition to X-ray films.
7.4.2.4.Chromogenic Read-Out Several examples for chromogenic read-out and densitometric quantification on peptide arrays are described. These are (1) precipitating substrates, e.g., nitroblue tetrazolium (NBT)/bromochloroindolyl phosphate (BCIP) catalyzed by alkaline phosphatase [269] or bromochloroindolyl-β-D-galactopyranoside catalyzed by β-galactosidase [155]; (2) metal ion detection with chromogenic chelators such as nickeldimethylglyoxime or as metal sulfides [269, 270, 494]; (3) heme [441] or metal ion [495] binding to four-helix bundle protein arrays; (4) binding of dye-coupled peptides to protein domain arrays [548]; and (5) chemical transformation such as crosslinking of advanced glycation end products (AGEs; [377]) or chemical ligation with dye-labeled aldehydes [538]. No expensive imager system is required for this type of read-out. Visual inspection is sufficient for quantification and documentation only requires a normal scanner. However, the dynamic range and sensitivity are far worse than, say, chemoluminescence or radioactivity. Another disadvantage is that regeneration of arrays on porous membranes treated with precipitating substrates, for example alkaline phosphatase or reagents for metal ion detection, is rather difficult since traces may be retained in the membrane structure.
7.4.2.5.Label-Free Read-Out Label-free read-out systems are the ultimate goal for screening peptide arrays. The screening molecule needs not to be modified, which is usually very tedious and may affect the biological activity of the analyte. Moreover, this excludes artifacts associated with the detection molecule, e.g. secondary antibodies or fluorescent markers, as discussed above. The most important label-free read-out systems are surface plasmon resonance (SPR), which can record kinetic data of the binding event [218, 581], mass spectrometry (MS), which can even identify a certain molecule out of a crude mixture, e.g., a cell lysate, with high sensitivity [278, 532], and atomic force microscopy (AFM) resulting in a three-dimensional image of the screening molecule bound to the array. So far, these technologies have mostly been used for DNA, protein, or small molecule arrays but it is just a question of time until they are used to a similar extend with peptide arrays.
Analysis of antibody-peptide interactions by SPR imaging using a peptide microarray was shown for interactions of the FLAG epitope and several of its variants with the anti-FLAG mab M2 [581]. First, a self-assembled monolayer (SAM) was prepared on a gold film and chemically modified to immobilize peptides. The FLAG peptide, three single substitution variants and an HA tag peptide as a negative control were tethered to this SAM as a “peptide-line” array using a microfluidics system. A second perpendicular microfluidics system was then used to deliver the analyte mab M2. In another study phosphorylation of a peptide substrate by the tyrosine kinase c-Src was measured by SPR [218].
PEPTIDE ARRAYS IN PROTEOMICS AND DRUG DISCOVERY |
221 |
The peptide was chemically immobilized on a SAM chip and phosphorylated by c-Src. Subsequently, phosphorylation was detected by SPR measurement of an anti-phospho-tyrosine antibody. Although in these studies only a few peptides were analyzed simultaneously, which does not really reach far beyond the capacity of standard SPR techniques as commercially supplied by Biacore for example, this read-out method has a huge potential for peptide microarrays.
Array-based read-out methods employing MS were first introduced as a robust method in the ProteinChip R Array Technology by the company Ciphergen [585]. Peptides or other molecules are immobilized on carriers with eight different positions. Subsequently, the chip is probed with either pure molecules or crude mixtures such as cell lysates, serum, or urine. Molecules that bind to the peptides or other molecules immobilized on the chip are retained during automated washing steps and are detected by surface-enhanced laser desorption/ionization time-of-flight mass spectrometry (SELDI-TOFMS). This technology is not reviewed further here because it does not fit the scope of peptide arrays with respect to the number of peptides analyzed and the degree of potential miniaturization. The combination of mass spectrometry and array techniques in the strict sense has already been successfully applied in the field of protein arrays [54] and might similarly be used for peptide arrays in the future. In preliminary studies Su and Mrksich immobilized the peptide Ac-IYAAPKKKC-NH2 on self-assembled monolayers presenting ethylene glycol chains with maleimide functions [532, 533]. However, only the success of immobilization was shown by MALDI-TOF MS. No further assays or detection were carried out by mass spectrometry. The application of mass spectrometry to identify binding partners from peptide arrays is described in a patent application [278]. A peptide scan derived from the intracellular domain of the erythropoietin (EPO) receptor was incubated with cell lysates from EPO stimulated and unstimulated cells as well as different purified signal transduction proteins. Subsequently, tryptic digestion, mass spectrometry, and databank analysis revealed the identity of the peptide-bound proteins.
Atomic force microscopy has not been used with peptide arrays so far, however proofof concept has been confirmed with protein arrays (e.g., [64, 245, 373]). AFM was used as a read-out for peptide arrays in the broader sense in a study visualizing transmembrane peptides in planar phosphatidylcholine bilayers that were prepared via fusion of vesicles with a solid substrate [471]. However, there was no peptide sequence diversity and no clear-cut spatial addressability.
A very elegant label-free read-out method uses conventional compact disc technology [282]. Molecules, meaning also peptides, are immobilized on the polycarbonate surface of a CD. Read-out of the reflective metalized layer underneath by a standard polarized infrared laser as used in CD players is disrupted by binding of bulky ligands to the surface and leads to errors in the binary signal. Proof-of-concept was shown for streptavidin binding to immobilized biotin but the strategy should also be applicable for peptide arrays.
7.5. APPLICATIONS OF PEPTIDE ARRAYS
In this Section the main applications for peptide arrays are described in principal and illustrated by selected outstanding examples. In most of these examples the peptide arrays were prepared by the SPOT method. This is mainly because publications based on this
222 |
ULRICH REINEKE, JENS SCHNEIDER-MERGENER AND MIKE SCHUTKOWSKI |
technology dominate the literature, since the first full publication describing this synthesis concept dates back more than a decade now and such array preparation is not only simple but requires no specialized equipment. Most applications illustrated here can be applied in principle to all types of peptide arrays described in Section 7.2. For a comprehensive overview of most of the publications in the field please refer to Table 7.4 in Section 7.6.
7.5.1. Antibodies
The most frequent application of peptide arrays described so far is the mapping of antibody epitopes. This can be ascribed to the fact that antibody-antigen interactions are often used as model systems for the evaluation of novel peptide library techniques, for several reasons: (1) usually antibodies bind to their antigens with high affinity and specificity (disregarding cross-reactivity). (2) Many antibodies raised against proteins bind to linear epitopes, unlike other protein-protein interactions where this is only an exception (see 7.5.2.2.). (3) Antibodies can be easily detected using commercially available enzymeor fluorescence dye-coupled secondary antibodies. (4) Antibodies are stable and easy to handle, and (5) state-of-the-art techniques allow rapid and cheap preparation of sufficient antibody amounts. Although antibody-antigen interactions are discussed separately in this Section many characteristics described here are also valid for protein-protein interactions in general (see 7.5.2.).
7.5.1.1. Monoclonal Antibody Epitope Mapping: Linear Epitopes In linear epitopes (also referred to as continuous or contiguous epitopes) [24] the key amino acids mediating antibody contacts are located within one part of the antigen’s primary structure, usually a region not exceeding 15 amino acids in length. Peptides covering these sequences have affinities to the antibody within the range shown by the entire antigen.
Three-dimensional structures of antibody-antigen complexes obtained from X-ray crystallography reveal relatively large contact surfaces in a range between 500 and 1000 Å2 with more than 15 amino acids in contact with the binding partner. This led to the definition of the “structural epitope” comprising all contact residues as observed in the complex structure without considering their energetic contribution. On the other hand, extensive sitedirected mutagenesis studies have shown that only a few residues effectively contribute to the binding free energy. These residues are summarized as defining the “energetic epitope” building or “hot spot of binding” [108]. Here, it should be pointed out that linear peptide epitopes identified by protein sequence-derived peptide scans comprise the amino acids of the energetic epitope, as well as a few linking residues, rather than the structural epitope.
Table 7.4 summarizes the publications describing mapping and characterization of linear antibody epitopes. The peptide arrays used include peptide scans, amino acids scans, substitutional analyses, truncation libraries, deletion libraries, cyclization scans, all types of combinatorial libraries, and randomly generated libraries of single peptides. Today, experiments to identify and characterize linear antibody epitopes are standard techniques widely applied even in non-specialized laboratories.
As listed in Table 7.4 most of the peptide array applications for mapping antibody epitopes are based on the SPOT synthesis technique due to its simplicity and robustness. However, the earliest publication describing antibody binding to arrays of short linear peptides, in 1991, utilized light-directed spatially addressable peptide synthesis [151]. Furthermore,