


Фізіологія сільськогосподарських тварин
Якщо парціальний тиск кисню в альвеолярному повітрі становить близько 100 мл рт. ст., то кількість дорівнює:
0 об’єму на 1 л крові,
або 0,32 об.%.
Як бачимо, кількість кисню, розчиненого у плазмі крові, незначна, але саме ця його частина відіграє важливу роль у дифузії та обміні цього газу між еритроцитами і тканинами. За звичайних умов кількість цього кисню постійна, бо скільки його проникає у тканини, стільки ж водночас надходить у плазму крові з еритроцитів.
Аналогічно визначають кількість вуглекислого газу, розчинного у плазмі крові. На рис. 42 відображено стан вуглекислого газу у венозній та артеріальній крові.
ТРАНСПОРТ КИСНЮ КРОВ’Ю
Кисень транспортується кров’ю переважно у хімічно зв’язаному стані. У легенях кисень, який шляхом дифузії надійшов у плазму, швидко переміщується до еритроцитів, де з’єднується з гемоглобіном. Один грам гемоглобіну може зв’язати 1,34 мл3 кисню. Сполука кисню з гемоглобіном — оксигемоглобін — нестійка.
Нb + О2 НbO2.
|
|
|
Таблиця 14 |
|
Киснева ємкість крові у сільськогосподарських тварин |
||||
|
|
|
|
|
Тварина |
Киснева ємкість, |
Тварина |
Киснева ємкість, |
|
% |
% |
|||
|
|
|||
Корова |
15,4 |
Кінь |
14,9 |
|
Вівця |
16,9 |
Кріль |
16,0 |
|
Коза |
14,2 |
Курка |
15,0 |
|
Свиня |
17,8 |
Качка |
15,4 |
|
Вона має чотири атоми заліза, кожний з яких утримує одну молекулу кисню. Знаючи вміст гемоглобіну у крові тварин (в середньому 14 г %), можна вирахувати кисневу ємкість крові. Вона характери-
106
Розділ 3. Дихання
зується максимальною кількістю кисню, яка може бути зв’язана 100 мл крові, за умови, якщо увесь гемоглобін перетвориться в оксигемоглобін. Так, 1,34 14=18,7, або близько 19% (об. доля), з урахуванням кисню, розчиненого у плазмі крові. Кисневу ємкість крові у сільськогосподарських тварин подано у таблиці 14.
|
Фіксація кисню гемоглобіном чи дисо- |
|
|
ціація оксигемоглобіну залежить в осно- |
|
|
вному від напруження цього газу у кро- |
|
|
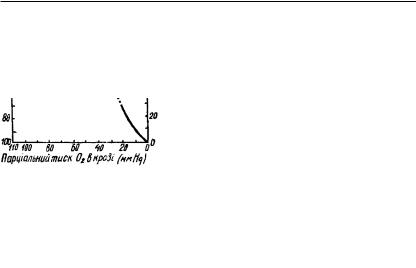
ві. На кривій Баркрофта (рис. 43) видно, |
|
|
що при збільшенні напруження кисень |
|
|
перетворюється в оксигемоглобін, але |
|
|
прямо пропорційної залежності ут- |
|
|
ворення оксигемоглобіну від напружен- |
|
|
ня газу не спостерігається. Так, при на- |
|
|
пруженні кисню 30 мм рт. ст. 60% |
|
|
гемоглобіну перетворюється в оксиге- |
|
Рис. 43. Крива (Баркрофта) |
моглобін, а при напруженні 60 мм рт. |
|
ст. — 90%, тобто майже весь гемоглобін |
||
фіксації кисню гемоглобіном |
окислюється. Така залежність фіксації |
|
залежно від парціального |
||
тиску цього газу в крові |
кисню гемоглобіном гарантує нормаль- |
|
не постачання цього газу до тканин, на- |
||
|
віть в умовах, коли процентний вміст кисню у вдихуваному повітрі зменшиться у два рази. При парціальному тиску кисню в альвеолярному повітрі 100 мм рт. ст. тільки 96% гемоглобіну крові легень перетворюється в оксигемоглобін. Верхня частина кривої свідчить про фіксацію кисню гемоглобіном у легенях, нижня відображає дисоціацію оскигемоглобіну на рівні тканин, а середня — стан у венозній крові.
Встановлено, що при підвищенні температури фіксація кисню гемоглобіном знижується. Така залежність сприяє звільненню його від гемоглобіну у робочих органах, де температура завжди вища, ніж у легенях. Підвищення концентрації вуглекислого газу та зниження рН крові у тканинах також сприяють звільненню кисню і переходу його у міжклітинну рідину. Навпаки, зниження концентрації двоокису вуглецю, температури та кислотності крові у легенях зумовлює утворення оксигемоглобіну. Слід відзначити, що гемоглобін плода має більшу спорідненість з киснем, ніж гемоглобін матері. Завдяки
107

Фізіологія сільськогосподарських тварин
цій властивості, незважаючи на незначну різницю в напруженні цього газу у плаценті, кисень шляхом дифузії проникає у кров плода. Цим і пояснюється перебування плода в стані апное.
ТРАНСПОРТ ВУГЛЕКИСЛОГО ГАЗУ КРОВ’Ю
Вуглекислий газ утворюється на рівні клітин, тому у тканинах його найбільше (60–70 мм рт. ст.). У міжклітинній рідині та венозній крові концентрація газу зменшується (46 мм рт. ст.). За рахунок такої різниці (14–24 мм рт. ст.) він переміщується з тканин у венозну кров.
Вище було зазначено, що коефіцієнт абсорбції двоокису вуглецю значно більший (0,592), ніж коефіцієнт абсорбції кисню, тому цей газ легше розчиняється у плазмі крові, його кількість становить 2,7–3 мл у 100 мл крові. Отже, частина вуглекислого газу транспортується у фізично розчинному вигляді. Основна частина його переноситься у хімічно зв’язаному стані, а саме у формі НСО3. Ці іони є у плазмі крові, де вони зв’язані з іонами Nа+, і в еритроцинах у комплексі з К+.
Таким чином, вуглекислий газ транспортується венозмою кров’ю в чотирьох формах. Найбільше його переноситься у формі CO3HNa, менша кількість у вигляді СО3НК, ще менша — у комплексі з гемоглобіном (карбо-гемоглобін) і зовсім мало — у фізично розчинному стані. Всього у венозній крові до 60% (об. доля) вуглекислого газу. В еритроцитах він може з’єднуватись з водою, утворюючи вугільну кислоту
СО2+Н2О Н2СО3 Н++НСО3–.
Ця реакція каталізується ферментом еритроцитів — карбоангідразою, яка прискорює синтез вугільної кислоти чи її розпад залежно від напруження вуглекислого газу у крові. У тканинах, де концентрація двоокису вуглецю значна, карбоангідраза прискорює (у 150 разів) синтез вугільної кислоти, а в легенях, де вміст вуглекислого газу зменшується, цей же фермент сприяє дисоціації утвореної раніше вугільної кислоти. Каталізуюча дія карбоангідрази виявляється тільки в еритроцитах, бо у плазмі крові цього ферменту немає. Незважаючи на це в еритроцитах міститься всього 30–50% загальної кількості вугільної кислоти, що синтезується.
108

Розділ 3. Дихання
ТКАНИННЕ ДИХАННЯ
Внутрішнє, або тканинне, дихання відбувається на рівні клітин організму. Воно складається з таких процесів: обмін кисню між артеріальною кров’ю і міжклітинною рідиною, переміщення кисню до клітин, використання його у процесах окислення, утворення вуглекислого газу та його перехід з клітин у кров венозної частини капілярів. Окислення — це біологічний процес, що починається з відщеплення водню від речовини, яка окислюється. Цей процес відбувається за участю ферментів дегідраз. Після відщеплення водню речовина, збагачена киснем, розпадається на прості складові, що супроводжується звільненням енергії. Далі водень з’єднується з киснем, утворюючи воду. Вуглекислий газ утворюється внаслідок відщеплення вуглецю та кисню від карбоксильних груп за участю ферментів декарбоксилаз. У ссавців кількість утвореного вуглекислого газу становить 2–8 мл за 1 хв в 1 кг м’язів.
Згадані гази проникають через дві мембрани, а саме крізь шар ендотелію капілярів та клітинну мембрану. Механізм обміну газів у тканинах та легенях подібний. Це чисто фізичний процес обміну газів шляхом дифузії за рахунок різниці напруження їх в артеріальній крові та клітинах. Напруження кисню і вуглекислого газу у крові артеріальної частини капілярів становить відповідно 100 і 40, у клітинах — 0 і 60 мм рт. ст. Напруження вуглекислого газу у тканинах не постійне і залежить від активності органів. При максимальній активності напруження зростає до 70 мм рт. ст., а у стані відносного спокою не знижується за 50 мм рт. ст. Кисню у клітинах майже ніколи немає, бо він одразу ж витрачається на окисні процеси. У міжклітинній рідині його напруження становить близько 20 мм рт. ст.
Обмін газів у тканинах залежить не тільки від різниці їх напруження у крові та клітинах. Проникнення газу через тканину залежить також від швидкості руху крові у капілярах та константи дифузії газу. Це кількість газу (мл), що проникає за 1 хв через певну поверхню (см2) тканини на певну глибину (мм) при певному атмосферному тиску. Ця константа залежить від температури. За її величиною можна судити про споживання газу тканиною. Споживання кисню тканинами залежить від інтенсивності обмінних процесів у них, але в середньому тканини забирають тільки 40% кисню, що міститься у крові. Як уже зазначалось, артеріальна кров містить близько 20% (об. доля) цього
109