

Технология переработки / Технология Мяса
.pdfВпереднюю голяшку входят: нижняя половина лучевой и локтевой костей и кости запястья.
Грудной отруб: передняя граница - по линии отделения плечевого отруба; задняя - вдоль нижней трети тринадцатого ребра; верхняя - по линии, идущей от верхней трети первого к нижней трети последнего (тринадцатого) ребра; нижняя - вдоль реберной дуги до грудной кости.
Вотруб входят: грудная кость с хрящами и соответствующими частями тринадцати ребер.
Спинной отруб: передняя граница - по линии отделения лопаточного отруба; задняя - между одиннадцатым и двенадцатым ребрами; нижняя - по линии отделения грудного отруба. В отруб входят: часть пятого и шесть грудных позвонков, начиная с шестого по одиннадцатый, с соответствующими им частями ребер.
Пашина: границы отделения проходят по линии, идущей от коленного сустава до сочленения истинной и ложной частей тринадцатого ребра и далее вдоль реберной дуги до грудной кости.
Поясничный отруб: передняя граница - по линии отделения спинного отруба; задняя - между пятым и шестым поясничными позвонками; нижняя
—по линии отделения пашины и грудинки.
Впоясничный отруб входят два последних грудных позвонка с ребрами (без нижней трети) и пять поясничных позвонков.
Тазобедренный отруб: передняя граница - по линии отделения поясничного отруба; задняя - поперек берцовой кости на уровне нижней ее трети; нижняя - по линии отделения пашины.
Втазобедренный отруб входят: кости таза (подвздошная, лонная, седалищная), крестцовая кость, шестой поясничный и два хвостовых позвонка,
бедренная кость, коленная чашечка и верхние 2/3 берцовой кости.
Задняя голяшка: отделяется поперек берцовой кости на уровне нижней ее трети, с предварительным отделением ахиллова сухожилия в месте перехода его в мышечную ткань. В заднюю голяшку входят: нижняя треть берцовой кости, кости скакательного сустава и ахиллово сухожилие.
Разделка баранины и козлятины для розничной торговли
согласно ГОСТ 7596-81 Туши разделываются по следующей схеме:
163
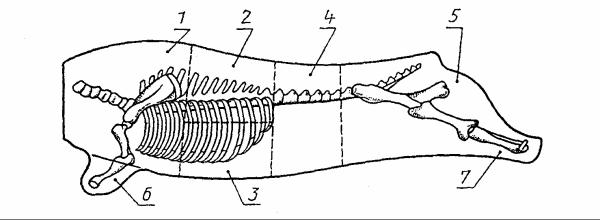
Отрубы подразделяются на сорта 1,2-й.
Ксорту 1 относятся: тазобедренный – 1, поясничный – 2, лопаточноспинной (включая грудинку и шею)-3.
Ксорту 2 относятся: зарез – 4, предплечье – 5, задняя голяшка – 6 Анатомические границы отделения отрубов должны проходить: Зарез: между вторым и третьим шейными позвонками.
В зарез входят два первых шейных позвонка.
Лопаточно-спинной отруб (включая грудинку и шею): передняя гра-
ница — по линии отделения зареза; задняя — между десятым и одиннадцатым ребрами перпендикулярно позвоночнику; нижняя — через плечелоктевой сустав.
Вотруб входят: пять шейных (с 3 по 7) позвонков, лопаточная и плечевая кости, десять грудных позвонков с соответствующими им ребрами и грудная кость с хрящами.
Предплечье: граница проходит через плече-локтевой сустав. В предплечье входят лучевая и локтевая кости и кости запястья.
Поясничный отруб: передняя граница — по линии отделения лопа- точно-спинного отруба; задняя — между пятым и шестым поясничными позвонками перпендикулярно позвоночнику.
Вотруб входят: три грудных позвонка и ребра (с 11 по 13), пять поясничных позвонков, часть пашины, а также почки с околопочечным жиром.
Тазобедренный отруб: передняя граница — по линии отделения поясничного отруба; задняя — через середину берцовой кости.
Вотруб входят: один поясничный и все хвостовые позвонки, кости таза (подвздошная, лонная, седалищная), крестцовая и бедренная кости, верхняя половина берцовой кости и часть пашины.
Задняя голяшка отделяется через середину берцовой кости с предварительным отделением ахиллова сухожилия в месте перехода его в мышечную ткань.
Взаднюю голяшку входят: нижняя половина берцовой кости, кости скакательного сустава и ахиллово сухожилие.
Разделка свинины для розничной торговли
согласно ГОСТ 7597-55 Свиные туши и полутуши разделываются на отдельные части
по нижеследующей схеме:
164
Наименование частей и сортность:
Ксорту 1 относятся: лопаточная – 1, спинная часть (корейка)-2, грудинка – 3, поясничная часть с пашиной -4, окорок - 5
Ксорту 2 относятся: предплечье (рулька) - 6, голяшка -7 Анатомические границы отделения отрубов должны проходить:
Лопаточная часть
Границы отделения проходят:
а) задняя - по прямой линии между пятым и шестым спинными позвонками с пересечением ребер;
б) нижняя — через плече-локтевой сустав.
Из костей в лопаточную часть входят: все 7 шейных позвонков, 5 первых спинных позвонков и соответствующие им ребра, лопатка с лопаточным хрящом, плечевая кость и передняя часть грудной кости.
Спинная часть (корейка)
Границы отделения проходят:
а) передняя — по линии отделения лопаточной части; б) задняя - впереди первого поясничного позвонка;
в) нижняя - поперек ребер, примерно, на половине их ширины.
Грудинка
Границы отделения проходят:
а) передняя - по линии отделения лопаточной части; б) задняя - за последним ребром; в) верхняя - по линии отделения спинной части.
Поясничная часть с пашиной
Границы отделения проходят:
а) передняя — по линии отделения спинной части и грудинки; б) задняя — по прямой линии, проходящей между последним и пред-
последним поясничными позвонками непосредственно впереди тазовой кости.
Окорок
Границы отделения проходят:
а) передняя — по линии отделения поясничной части с пашиной; б) задняя — по линии отделения голяшки.
Голяшка
Отделяется от окорока в поперечном направлении через верхнюю треть берцовых костей.
В заднюю голяшку входят две трети берцовых костей и скакательный сустав.
Предплечье (рулька)
Отделяется по прямой линии через плече-локтевой сустав. В рульку входят кости предплечья и запястный сустав.
165
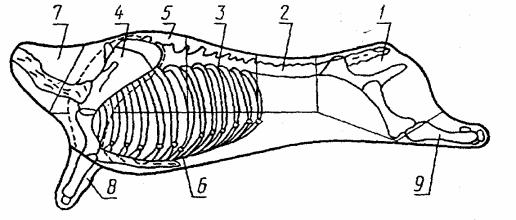
Разделка телятины для розничной торговли
согласно ГОСТ 23219-78 Туши разделываются по следующей схеме:
В соответствии с наименованиями отрубы подразделяются на 1,2,3 сор-
та
Ксорту 1 относятся: тазобедренный – 1, поясничный – 2, спинной – 3, лопаточный - 4, подплечный край -5.
Ксорту 2 относятся: грудной с пашиной – 6, шейный – 7.
Ксорту 3 относятся: предплечье -8, голень - 9
Анатомические границы отделения отрубов должны проходить: Шейный отруб - между пятым и шестым шейными позвонками. В шейный
отруб входятпятьшейных позвонков (с первого по пятый); Лопаточный отруб - от реберной части по фасциям; от грудной части путем
разреза мышечной ткани; от предплечья - через локтевой сустав. В лопаточный отруб входят лопаточная и плечевая кости;
Предплечье - через локтевой сустав. В отруб входят лучевая, локтевая кости и кости запястья;
Грудной с пашиной - по линии, проходящей в направлении от коленной чашечки к нижней трети последнего ребра и далее к середине первого ребра. В грудной отруб входит грудная кость с хрящами и соответствующими частями тринадцати ребер;
Подплечный край - передняя граница - по месту отделения шейного отруба; задняя - между седьмым и восьмым ребрами и соответствующими им позвонками; нижняя - по линии отделения грудинки. В подплечный край входят два шейных и семь первых грудных позвонков, с соответствующими им частями ребер;
Спинной отруб - передняя граница - по линии отделения подплечного края; задняя - между последним грудным и первым поясничным позвонками; нижняя - по линии отделения грудного отруба. В спинной отруб входят шесть грудных позвонков (с восьмого по тринадцатый) и соответствующие им части ребер;
Поясничный отруб - передняя граница - по линии отделения спинного отруба; задняя - между пятым и шестым поясничными позвонками; нижняя
166
— по линии отделения пашины. В поясничный отруб входят пять поясничных позвонков;
Тазобедренный отруб - передняя граница - по линии отделения поясничного отруба; нижняя - по линии отделения пашины; задняя - через коленный сустав. В тазобедренный отруб входят шестой поясничный и два хвостовых позвонка, кости таза (подвздошная, лонная, седалищная), крестцовая и бедренная кости, коленная чашечка;
Голень - через коленный сустав по направлению к ахиллову сухожилию. В голень входят берцовая кость и кости скакательного сустава.
167
Глава V. Изменения в мясе после убоя и при хранении. Характеристики мяса с признаками PSE и DFD. Генетически модифицированные продукты.
5.1Сущность послеубойных изменений в мясе. Автолиз. Созревание мяса. Предшественники вкуса и аромата
Классическое течение автолиза. Определяющим условием для фор-
мирования качества мяса являются уровень и характер развития автолитических процессов, начинающихся немедленно, после убоя животного. Автолиз
(или самораспад) - это прекращение обмена веществ в тканях в послеубойный период и переход обратимых биохимических процессов в необ-
ратимые под действием тканевых ферментов и микроорганизмов. Физи-
ко-химические, гистологические и органолептические изменения протекают в мясе после убоя животного в соответствии с основными этапами автолиза: парное мясо - посмертное окоченение - разрешение посмертного окоченения и созревание. Стадия послеубойного хранения мяса, характеризующаяся развитием процесса окоченения, сопровождается различными биохимическими процессами и изменениями в клеточной структуре:
распадом гликогена и образованием молочной кислоты, смещением рН от нейтральной точки в кислую сторону;
распадом высокоэнергетических фосфатов креатинфосфорной (КФ) и аденозинтрифосфорной (АТФ) кислот;
перераспределением ионов Са 2+ в клеточных структурах мышечного волокна;
ассоциацией актина и миозина в актомиозиновый комплекс; изменениями в составе и свойствах гемовых пигментов; началом процессов окисления жиров; изменением микроструктуры мышечного волокна.
Интенсивность гликолитических превращений в автолизирующей мышечной ткани различных видов животных определяется предубойным состоянием животного, функциональными особенностями соответствующих мышц и зависит от концентрации гликогена и активности многих ферментов, участвующих в гликолизе.
Сразу после убоя животного ткани находятся в расслабленном состоянии, мышечные волокна набухшие, прямолинейной или слегка волокнистой формы, соединительно-тканные волокна сокращены волнообразно. Это состояние характеризуется высоким значением растворимости фракции белков. Мясо в период до посмертного окоченения сохраняет запас гликогена и АТФ и основные мышечные белки - миозин и актин - в диссоциированном состоянии обладают хорошими гидрофильными свойствами, что обеспечивает их высокую водосвязывающую способность. Содержание АТФ в мышцах парных туш примерно такое же, как и в мышцах живых животных, что также способствует хорошей водосвязывающей способности. Так, парное мясо при
168
величине рН 5,9 поглощает в среднем 86% воды, а охлажденное (рН 5,4) - только 33%. Потеря водосвязывающей способности обусловлена в основном распадом АТФ и на одну треть понижением величины рН.
Преимущества парного мяса проявляются и в свойствах белков соединительной ткани. Развариваемость коллагена в первые часы после убоя животных достигает 20-30%) от его исходного содержания, а после хранения в течение 2 суток уменьшается до 14-18%.
Парное мясо характеризуется сравнительно большим содержанием ароматических веществ и летучих жирных кислот, от которых, в свою очередь, зависят его вкус и аромат. Оно обладает хорошо выраженными бактериостатическими свойствами по отношению ко многим видам бактерий, поэтому размножение микроорганизмов в нем заторможено.
Высокую водо - и жиросвязывающую способности, нежную консистенцию, максимальную развариваемость коллагена, стабильный цвет и аромат, минимальную микробную обсемененность парное мясо сохраняет в течение ограниченного времени (по данным разных исследователей - от 3 до 10 ч), т.е. до наступления посмертного окоченения. Параллельно с его наступлением теряются свойства парного мяса. По прошествии некоторого времени после убоя (3-4 ч) в тканях начинают преобладать процессы окоченения мышц с накоплением в тканях молочной кислоты и образованием актомиозинового комплекса. По мере развития этого процесса, мясо теряет свою эластичность и становится жестким, трудно поддается механической обработке (обвалке, резанию, жиловке). Максимум изменения прочностных свойств мяса совпадает с достижением максимального окоченения. К этому же моменту водосвязывающая способность уменьшается до минимума.
В процессе посмертного окоченения мышц происходят, преиму-
щественно, распад гликогена, КФ и АТФ, ассоциация актина и миозина в актомиозиновый комплекс и сокращение гидратации мышц.
Распад гликогена в первые часы автолиза идет, преимущественно, путем фосфоролиза, который постепенно замедляется к 24 ч после убоя, вследствие исчезновения АТФ и накопления молочной кислоты. К этому моменту рН мышечной ткани достигает величины 5,4-5,8 и приближается к изоэлектрической точке белков мышечного волокна (4,7-5,4). При участии лизосомальных ферментов усиливается амилолитический распад гликогена на фоне снижения величины рН. Уровень изменения этого показателя зависит от прижизненного содержания мышечного гликогена, составляющего в норме около
1%.
Содержание АТФ в мышечной ткани является совокупным результатом ее ресинтеза в процессе гликолиза и гидролитического распада под действием миозиновой АТФ-азы. АТФ гидролизуется с образованием аденозиндифосфорной кислоты (АДФ) и свободного неорганического фосфата, а освобождающаяся химическая энергия превращается в механическую энергию мышечного сокращения. Наряду с АТФ, сразу же после убоя распадается КФ.
Скорость развития окоченения зависит от количества АТФ, содержащегося в мускулах. При наличии в тканях резервов гликогена полный
169
распад АТФ не может произойти и мускул не переходит в состояние полного окоченения. Распад АТФ и КФ оказывает существенное влияние на состояние миофибриллярных белков, определяющих консистенцию мяса, его способность связывать воду, эмульгирующие и адгезионные свойства. Энергия распада АТФ стимулирует взаимодействие миозина с актином с образованием актомиозинового комплекса, в результате чего длина саркомеров уменьшается и происходит развитие мышечного окоченения. Указанное явление сопровождается увеличением механической прочности мышечных волокон, резким уменьшением гидратации белков миофибрилл и водосвязывающей способности мяса. Интервал между рН среды и изоэлектрической точкой белков мяса уменьшается, вследствие чего сокращается число ионизированных групп белков. Большая часть белков вообще переходит в изоэлектрическое состояние, что способствует их агрегации и снижению водосвязывающей способности мяса.
В ходе автолиза претерпевают изменения также белки соединительной ткани (коллаген, эластин). В состоянии окоченения они становятся менее лабильными, чем, непосредственно, после убоя. На стадии посмертного окоченения резко понижается развариваемость коллагена. При появлении первых признаков окоченения обнаруживаются многочисленные участки деформированных мышечных волокон и прилегающей к ним, соединительной ткани.
На стадии посмертного окоченения аромат и вкус мяса плохо выражены, наблюдается понижение гидролиза белков ферментами желудочнокишечного тракта. Время наступления и продолжительность окоченения зависят от состояния животного перед убоем, скорости охлаждения мяса и температуры хранения, вида животных и их упитанности. Так, при 0 ° С говядина
находится в состоянии окоченения 2 суток, |
а при 18 ° С - 1 сутки. Полное |
развитие окоченения говядины и баранины |
наступает через 18-24 ч, свини- |
ны - через 16-18 ч. |
|
В результате смещения рН в кислую среду увеличивается устойчивость мяса к действию гнилостных микроорганизмов, снижаются растворимость мышечных белков и уровень их гидратации, происходит набухание коллагена и повышается активность мышечных катепсинов, усиливающих гидролиз белков на более поздних стадиях автолиза, разрушается бикарбонатная система мышечной ткани, создаются условия для реакций цветообразования, интенсификации аромата и вкуса и т.д.
После завершения процесса окоченения происходит постепенное размяг-
чение мышечной ткани. Мясо становится нежным и сочным, приобретает соответствующие вкусовые и ароматические характеристики. Вещества, формирующие вкус и аромат мяса, являются продуктами распада белков и пептидов (глютаминовая кислота, треонин, серосодержащие аминокислоты и др.), нуклеотидов (инозинмонофосфорная кислота, инозин, гипоксантин, рибоза), углеводов (глюкоза, фруктоза, молочная и пировиноградная кислоты), липидов (низкомолекулярные жирные кислоты), а также и другие вещества (креатин, креатинин).
170
Формирование качества мяса при созревании обусловлено комплексом ферментативных и неферментативных процессов, вызывающих изменение состава и состояния основных компонентов мяса. При созревании начинаются частичная диссоциация актомиозина на актин и миозин и переход актомиозина из сокращенного в расслабленное состояние. При этом увеличивается число гидрофильных центров миофибриллярных белков, что обусловливает рост водосвязывающей способности мышечной ткани. Повышение протеолитической активности в мышцах происходит вследствие высвобождения из лизосом ферментов - катепсинов, которые наиболее заметно воздействуют на белки саркоплазмы. Вместе с тем, ограниченному протеолизу подвергаются и миофибриллярные белки. Расщепление небольшого количества пептидных связей в этих белках достаточно для разрыхления структур и увеличения нежности мышечной ткани.
В изменении нежности мяса важную роль играют количество и состояние компонентов соединительной ткани. Лабильность компонентов соединительной ткани увеличивается в период созревания мяса, когда из лизосом высвобождаются гидролитические ферменты. В результате образуются растворимые продукты распада коллагена, повышается растворимость основного вещества соединительной ткани и коллаген легче разваривается. Воздействие кислот, образующихся в процессе созревания мяса приводит к некоторому разрыхлению коллагеновых пучков, ослаблению межмолекулярных поперечных связей и набуханию коллагена, что также способствует получению более нежного мяса.
В процессе созревания различные компоненты мяса претерпевают неодинаковую степень превращений, характерным образом влияющие на изменение нежности. Так, химические продукты, образующиеся при послеубойных изменениях жировой ткани, ухудшают качество жира, и по качественным показателям лучшим считается жир, непосредственно, после убоя животного. Поэтому при равных условиях созревания различных отрубов мяса одного и того же животного, а также одинаковых отрубов разных животных нежность его оказывается различной.
Продолжительность созревания мяса зависит от температуры, упитан-
ности и возраста животных, вида мяса. При повышенной температуре мясо созревает быстрее; упитанных животных - дольше, молодняка - быстрее; говядина при 0° С созревает 10-14 суток, при 8-10° С -6 суток, а при 16-18 ° С - 4 суток; баранина, свинина созревают быстрее: при 0 ° С соответственно 8 и 10 суток.
Таким образом, в процессе созревания, мясо приобретает хорошо выраженный аромат и вкус, становится мягким и сочным, более влагоемким и более доступным действию пищеварительных ферментов, имеет более высокую пищевую ценность по сравнению с мясом в состоянии посмертного окоченения.
Следует отметить, что при хранении созревшего мяса в асептических условиях при низких положительных температурах под влиянием внутриклеточных ферментов автолитические процессы в мясе продолжаются. На ста-
171
дии глубокого автолиза происходит разрыв пептидных связей белковых частиц и распад белковых веществ. Причем, некоторые из образовавшихся при распаде белков вещества, обладают токсическими свойствами. Под действием липаз происходит интенсивный гидролитический распад жиров. При распаде белковых веществ происходит разрушение морфологических структурных элементов мышечной ткани. Изменяются окраска и вкус мяса: оно приобретает коричневый оттенок и неприятный кислый вкус. На этапе глубокого автолиза мясо может даже стать непригодным для употребления в пищу. Однако, в практике промышленной переработки мяса глубокий автолиз, практически, не встречается, так как микробиальная порча наступает раньше.
Как очевидно из сказанного, посмертные изменения, обусловленные содержанием молочной кислоты и величиной рН, имеют важное практическое значение и оказывают существенное влияние на качество мяса, его пищевую ценность и технологические характеристики (потери мяса в процессе обработки, изменение нежности мяса, водосвязывающая способность, аромат и вкус, переваримость под действием пищеварительных ферментов, устойчивость к микробиологическим процессам, сроки хранения, уровень потерь воды при тепловой обработке, количество мясного сока, выделяющегося при размораживании). Поэтому следует определять целесообразность использования мяса на реализацию, хранение или промышленную переработку, в зависимости от уровня развития автолитических процессов.
Накопление веществ, обусловливающих запах и вкус. Свежее мясо име-
ет незначительные специфические вкус и запах. В процессе созревания в результате автолитических превращений белков, липидов, углеводов и других компонентов образуются низкомолекулярные вещества, формирующие запах и вкус мяса.
Однако, отчетливо выраженные вкус и запах появляются лишь после тепловой обработки мяса, следовательно, в процессе автолиза в мясе образуются и накапливаются предшественники веществ, формирующие запах и вкус при кулинарной обработке.
Характерный вкус мяса и мясного бульона зависит от содержания в составе мышечной ткани глютаминовой кислоты, которая, так же как и ее соли
– глютаминаты, обладают специфическим вкусом мясного бульона.
Аромат и вкус пищевых продуктов, возникающие при их тепловой обработке, находятся в прямой связи с реакцией меланоидинообразования, резко ускоряющейся при нагреве. Эта реакция протекает с участием аминокислот и моносахаридов с образованием летучих карбонильных соединений.
Изменение аромата и вкуса мяса, в процессе его созревания, связано также с накоплением низкомолекулярных летучих жирных кислот, образующихся в результате гидролитического расщепления липидов кишечного волокна под действием липазы. Известно, что жирные кислоты с числом углеродных атомов менее 12 обладают более или менее выраженными ароматом и вкусом. Различие в жирнокислотном составе липидов мышечного волокна разных животных придает специфичность оттенкам аромата и вкуса различных видов мяса.
172