

Глюкоза Этиловый
спирт
2. Молочнокислое брожение идет по уравнению:
СбН12Об 2 СН3СНОН—СООН
3. Маслянокислое брожение:
СбН120б — СН3—СН2—СН2—СООН+2СО2+2Н2
Масляная кислота
Первые этапы при различных видах брожения идут аналогично вышеописанному анаэробному распаду глюкозы при гликолизе до образования пировиноградной кислоты. Так же, как это было рассмотрено выше для животных тканей, в микроорганизмах (дрожжах, бактериях) происходит прежде всего фосфорилирование глюкозы с образованием глюкозо-6-фосфат, а затем фруктозо-1,6-дифосфат. Дифосфат фруктозы также распадается на две молекулы глицеринового альдегида, а последний через ряд стадий дает пировиноградную кислоту.
Анаэробный распад углеводов в дрожжах (спиртовое брожение) приводит к образованию этилового спирта. При этом брожении пировиноградная кислота на первом этапе подвергается де-карбоксилированию под действием пируватдекарбоксилазы, кофер-ментом которой является тиаминдифосфат (тиаминпирофосфат). В результате образуется СО2 и уксусный альдегид, последний затем при участии восстановленной алкогольдегидрогеназы восстанавливается в спирт.
1. СН3—СО—СООН С02+СН3—С==0
Пировиноградная пируватдекарбок-
кислота силаза H
Уксусный
альдегид
2. СНз-С==O НАДН+Н+
 алкогольдегидрогеназа
С2Н5ОН
+ НАД+
алкогольдегидрогеназа
С2Н5ОН
+ НАД+
Н
На процессе спиртового брожения основано виноделие и спирто-водочное производство.
Молочнокислое брожение идет при участии молочнокислых бактерий. Восстановленная дегидрогеназа (лактатдегидоогеназа) передает свой водород пировиноградной кислоте, которая восстанавливается в молочную кислоту.
лактатдегидро-
С
 Н3-СО-СООН
+ 2Н
СН3СH-
ОН –
СООН
Н3-СО-СООН
+ 2Н
СН3СH-
ОН –
СООН
геназа Молочная кислота
Молочнокислое брожение играет большую роль при производстве молочнокислых продуктов (кефира, простокваши и др.), при квашении капусты, огурцов, при силосовании кормов. Маслянокислое брожение вызывается маслянокислыми бактериями, большинство из которых являются анаэробами. Образование масляной кислоты также идет через пировиноградную кислоту.
Аэробное окисление углеводов.
Распад углеводов в аэробных условиях может идти прямым (aпотомическим или пентозным) путем и непрямым (дихотомическим) путем.
Дихотомическое (греч. dicha - на две части, tome-сечение) окисление углеводов идет по уравнению:
C6H12O6+6O2 6 СО2+б Н2О+686 ккал
Этот путь является основным в образовании энергии. Первые этапы этого пути совпадают с анаэробным окислением глюкозы. Расхождение путей начинается на стадии образования пировиноградной кислоты, которая в животных тканях декарбоксилируется окислительным путем.
Первоначально предполагали, что пировиноградная кислота подвергается окислительному декарбоксилированию с образованием уксусной кислоты по уравнению: СН3—СО—СООН + 1/2 O2 СН3СООН+СО2. Потом было установлено, что уксусная кислота не является промежуточным продуктом при .декарбоксилировании пировиноградной кислоты, и возникло представление, что уксусная кислота существует в «активной» форме. Вскоре было показано, что для утилизации пирувата необходим коэнизм А (КоА). В химическом отношении КоА представляет нуклеотид, в состав которого входит аденозин-31, 51-дифосфат, фосфат, пантотеновая кислота и тиоэтиламин. Коэнзим А участвует в переносе остатка уксусной кислоты — ацетильного радикала (CH3CO-) также и других кислотных (ацильных) радикалов.
Окислительное декарбоксилирование пирувата — процеcc многоступенчатый, осуществляется сложной ферментативной системой, в состав которой помимо пируватдекарбоксилазы, тиаминдифосфата и коэнзима А входят дегидрогеназы с коферментом НАД+, и ФАД, липоевая кислота и ионы магния. В результате окисления пировиноградной кислоты образуются молекула aцетил-КоА («активной» формы уксусной кислоты), два атома водорода (в виде НАДН+Н+ ) и молекула СО2.
O
СН3-СО-СООН+НS-КоА+НАД+СН3--С~S-~КоА+С02+НАДН+ H
Ацетил-КоА
Следующий этап непрямого аэробного окисления глюкозы характеризуется полным окислением ацетил-КоА в цикле Кребса до СО2 и Н2О.
ЦИКЛ ДИ- и ТРИКАРБОНОВЫХ КИСЛОТ (КРЕБСА).
Исследования показали, что дальнейшее окисление ацетил-КоА возможно лишь в присутствии небольших количеств какой-либо ди-карбоновой кислоты. Оказалось, что в начале ацетил-коэнзима А конденсируется с щавелевоуксусной кислотой (СООН—СН2— СО—СООН) с образованием лимонной (трикарбоновой) кислоты. Лимонная кислота является первым продуктом цикла Кребса, поэтому этот цикл иногда называют лимоннокислым.
Образовавшаяся
лимонная кислота подвергается далее
ряду сложных превращений. И прежде
всего, дегидратированию с образованием
цис-аконитовой кислоты. Последняя
присоединяет молекулу воды и переходит
в изолимонную кислоту. Изолимонная
кислота подвергается дегидрированию
и превращается в щавелево-янтарную,
которая декарбоксилируется с образованием
-кетоглютаровой.-Кетоглютаровая
кислота подвергается окислительному
декарбоксилированию и одновременно
дегидрируется, переходя в сукцинил-КоА.
Затем сукцинил-КоА превращается в
янтарную кислоту. Янтарная кислота
дегидрируется, превращаясь в фумаровую.
Фумаровая переходит в яблочную, а из
яблочной при ее дегидрировании образуется
щавелевоуксусная. На этом цикл замыкается.
Многие реакции цикла Кребса легко
обратимы. Ферменты, катализирующие
реакции цикла Кребса с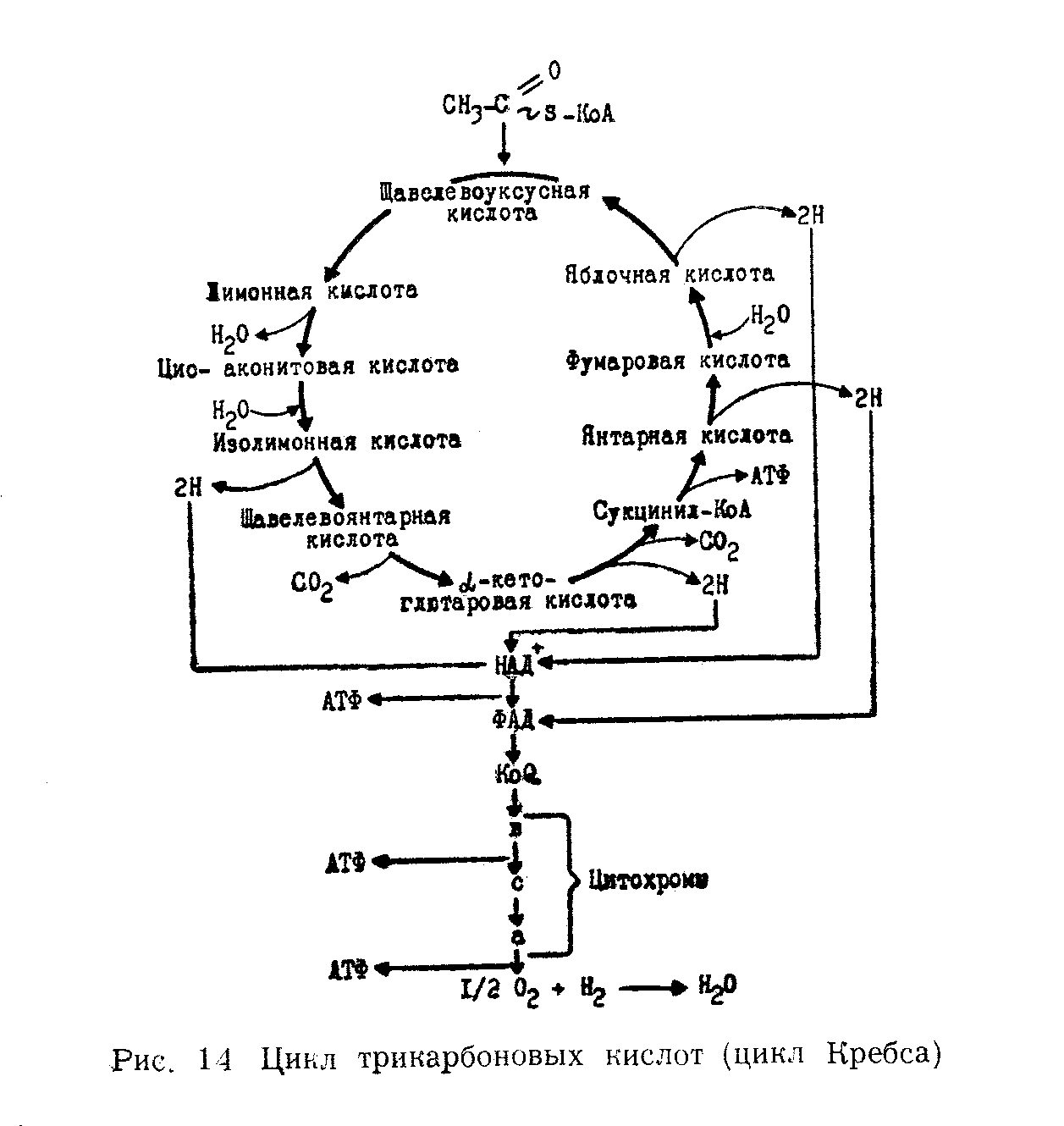 осредоточены
в митохондриях. Последовательность
реакций цикла Кребса последовательно
изображена на суммарной схеме (рис. 7.3)
.
осредоточены
в митохондриях. Последовательность
реакций цикла Кребса последовательно
изображена на суммарной схеме (рис. 7.3)
.
Из приведенной схемы видно, что в цикле Кребса в результате реакций дегидрирования образуется 4 пары водородных атомов и 2 молекулы СО2. Освободившийся в ходе процесса окисления водород поступает в цепь биологического окисления и в конечном итоге окисляется молекулярным кислородом с образованием воды и выделением энергии.
При окислении в цикле ди- и трикарбоновых кислот одной молекулы ацетил-КоА образуется 12 молекул АТФ, из которых одиннадцать возникает путем окислительного фосфорилирования, а одна при субстратном фосфорилировании (при превращении сукцинил-КоА в янтарную кислоту).При окислении большинства субстратов в цепи биологического окисления происходит образование 3-х молекул АТФ, тогда как окисление некоторых из них (например, в случае янтарной кислоты) дает 2 молекулы АТФ. Энергетический баланс анаэробного и аэробного окисления глюкозы представляет следующую картину.
1. Две молекулы АТФ — это чистый прирост АТФ при превращениях глюкозы до пировиноградной кислоты в анаэробной фазе.
Глюкоза+2 НАД+ +2 АДФ+2 Фн- 2 Пируват+2 НАДН+2 АТФ
2. Четыре молекулы АТФ образуются в результате окисления двух молекул НАДН, возникших при дегидрировании двух молекул 3-фосфоглицеринового альдегида (рис. 13). В связи с тем, что эти две молекулы НАДН являются цитоплазматическими, то отдаваемые ими электроны могут включиться в митохондриальную цепь биологического окисления не прямым путем, а с помощью так называемого челночного механизма. Суть этого механизма состоит в том, что сначала цитоплазматический НАДН реагирует с фосфодиоксиацетоном и образует глицерол-3-фосфат.
Фосфодиоксиацетон+НАДН Глицерол-3-фосфат+НАД+
Г'лицерол-3-фосфат легко проникает через митохондриальную мембрану и окисляется с участием флавинзависимой дегидрогеназы в фосфодиоксиацетон, причем простетическая флавиновая группа восстанавливается.
Глицерол-3-фосфат+Фл. пр. Фосфодиоксиацетон+
4-фл. пр. Н2. Фосфодиоксиацетон выходит из митохондрий, а восстановленный флавопротеид (Фл. пр. Н2) передает приобретенные электроны в цепь биологического окисления, обеспечивая окислительное фосфорилирование только двух молекул АДФ.
3. Шесть молекул АТФ возникают в процессе окислительного де-карбоксилирования двух молекул пировиноградной кислоты, образовавшихся в анаэробной .фазе из одной молекулы глюкозы.
4. При полном окислении двух молекул ацетил-КоА в цикле Кребса возникает 24 молекулы АТФ. В итоге полного аэробного окисления одной молекулы глюкозы синтезируется 36 молекул ЛТФ. При анаэробном гликолизе (брожении) на одну молекулу глюкозы образуется всего 2 молекулы АТФ. Таким образом, «выход» энергии, запасаемой в виде АТФ при кислородном распаде глюкозы, в 18 раз больше, чем при анаэробном.
ПЕНТОЗО-ФОСФАТНЫЙ ПУТЬ ОКИСЛЕНИЯ УГЛЕВОДОВ.
Существует еще один путь аэробного окисления углеводов, который имеет значение лишь в некоторых органах и тканях (жировой ткани, печени, эритроцитах, молочной железе, коре надпочечников, половых железах). Этот путь называется пентозофосфатным (пентозным, прямым или гексозомонофосфатным).
В отличие от непрямого (гексозодифосфатного) пути окисления углеводов в пентозном цикле не проходит стадии образования фруктозе-16-дифосфата, фосфотриоз и т. д., т. е. не происходит фосфорилирования глюкозомонофосфата. Последний в пентозном цикле подвергается прямому окислению с образованием в конечном итоге СО2 и пентозофосфата.
Значение пентозного цикла состоит, во-первых, в том, что является основным поставщиком восстановленного НАДФ + (НАДФН), необходимого для разнообразных синтетических процессов. Во-вторых, этот цикл обеспечивает организм пентозами. В третьих, энергетическая ценность цикла весьма велика — равна 36 мол АТФ.
7.5. ФОТОСИНТЕЗ
Фотосинтез - это синтез органических веществ (прежде всего углеводов) из углекислого газа и воды, происходящий за счет энергии света. Фотосинтез является основным источником образования органических веществ на Земле и единственным источником кислорода.
Процесс фотосинтеза в растениях можно выразить следующим
свет
суммарным уравнением: 6С02+6Н20 С6 Н 12О6+6 O2
хлорофилл
фотосинтез происходит внутри специализированных органелл- пластидах (хлоропластах), которые содержат в себе хлорофилл.
Хлорофилл — сложное гетероциклическое соединение, содержащее четыре перрольных кольца, образующих порфириновое ядро. Перрольные кольца связаны двумя основными и двумя дополнительными валентностями с атомом магния. Хлорофилл благодаря наличию в его структуре сопряжённых двойных связей способен поглощать световую энергию, переходя в активное состояние. Аккумулирование солнечной энергии хлорофиллом является начальным этапом фотосинтеза, который можно изобразить следующим образом:
Хл + hv — Хл+
Хлоро- Квант Хлорофилл
филл света возбужденный
На втором этапе фотосинтеза возбужденный хлорофилл, обогащенный энергией за счет поглощения светового фотона, отдаёт энергию на разложение воды (фотолиз воды). В результате образуются активные радикалы Н и ОН. Радикалы ОН дают перекись (ОН)2, распад которой приводит к образованию кислорода.
Хл+ +Н20 — Хл+Н+ОН
ОН (ОН)2 —— H2O+1/2O2
Следующий (третий) этап состоит в переносе водорода воды
через цепь переносчиков на НАДФ с образованием НАДФН. Одновременно происходит за счет переноса электронов, индуцируемого светом, синтез АТФ из АДФ и неорганического фосфата, т. е так называемое фотосинтетическое фосфорилирование.
Разобранные выше этапы являются световой фазой фотосинтеза. В реакциях этой фазы принимает участие поглощенная хлорофиллом световая энергия. Световая фаза приводит к образованию молекулярного кислорода, НАДФН и АТФ.
Следующая фаза фотосинтеза — темновая. Реакции в этой фазе происходят без участия света. В темновой фазе фотосинтеза НАДФН и АТФ используются для восстановления углекислого газа до углевода.