

Глава 4. Ферменты
4.1.Общее понятие о ферментах. Иммобилизованные ферменты
Ферменты, или энзимы - это биологические катализаторы, образующиеся и функционирующие во всех живых организмах. Из приведенных названий в литературе на русском языке принят термин “ферменты”, а процессы, происходящие с участием этих катализаторов - ферментативными.
Вещество, превращение которого катализирует фермент, получило название с у б с т р а т.
Ферменты являются важнейшими компонентами клетки, они тесным образом связаны с разнообразными процессами жизнедеятельности, их роль как биокатализаторов биохимических превращений подобна роли катализаторов в других химических реакциях. По определению И.П.Павлова “ферменты есть... первый акт жизнедеятельности... они... возбудители всех химических превращений... основной пункт, центр тяжести физиолого-химического знания”.
Ферменты идеально приспособлены для работы в живой клетке, но после выделения из клетки они не теряют свои каталитические свойства. На этом основано практическое применение ферментов в химической, пищевой, легкой и фармацевтической промышленности.
Принцип связывания ферментов с различными структурами клетки в настоящее время используют в биотехнологии. При этом ферменты прикрепляют (иммобилизуют) к поверхности какого-либо твердого носителя (целлюлоза и ее производные, полиакриламид, пористое стекло, нейлон, алюмосиликаты и др.), что позволяет не только сохранить их каталитические свойства, но и повысить стабильность. Такие ферменты получили название и м м о б и л и з о в а н н ы х.
Иммобилизованные ферменты обладают рядом преимуществ по сравнению с природными предшественниками: во-первых, их можно легко отделить от реакционной среды и использовать повторно; во-вторых, процесс можно вести непрерывно (в проточных колоннах) и, изменяя скорость потока, регулировать скорость каталитической реакции и выход продукта. Иммобилизованные ферменты успешно используют для получения глюкозы из крахмала, получения глюко-фруктозного сиропа и в ряде других крупнотоннажных производств.
4.2. Химическая природа и строение ферментов. Активный центр ферментов
Установлено, что все известные в настоящее время ферменты представляют собой белки.
Ферменты обладают теми же физико-химическими свойствами, что и белки. Их молекулярная масса колеблется от десятков тысяч до нескольких миллионов. По форме молекул ферменты относятся к глобулярным белкам.
Все ферменты подразделяют на две большие группы: о д н о к о м п о н е н т н ы е и д в у х к о м п о н е н т н ы е. К первой группе относятферменты, состоящие только из белка, а ко второй - состоящие из белка и связанной с ним небелковой части (активная группа или кофактор). Белковая часть двухкомпонентного фермента носит название а п о ф е р м е н т, небелковая часть - п р о с т е т и ч е с к а я группа или кофермент, а молекула в целом - х о л о ф е р м е н т.
Прочность связи между белковой и небелковой частями у различных ферментов различна. В связи с этим небелковую часть, сравнительно прочно связанную с апоферментом называют п р о с т е т и ч е с к а я группа, анебелковую часть, сравнительно легко удаляющуюся через полупроницаемую мембрану при диализе - к о ф е р м е н т.
В качестве кофакторов двухкомпонентных ферментов может функционировать значительное число органических и неорганических веществ. Из органических соединений функцию кофакторов выполняют многие витамины, нуклеотиды (ФМН и др.), динуклеотиды (НАД, НАДФ, ФАД), железопорфирины (гем и гематин), липоевая кислота, и другие соединения.
Из неорганических веществ функцию кофакторов выполняют ионы различных металлов: цинка, меди, железа, молибдена, никеля, марганца, магния, кальция и др. В одних ферментах металлы бывают довольно прочно связаны с белком и не отделяются от него в процессе очистки. В других ферментах металл непрочно связан с белком и легко отделяется от него в процессе очистки.
А к т и в н ы й ц е н т р. Известно, что размеры ферментов намного превышают размеры субстратов или функциональных групп, на которые они действуют. Это дало основание предполагать, что субстрат соединяется не со всей молекулой фермента, а с отдельным его участком, получившим название “а к т и в н ы й ц е н т р”, т.е. та область фермента, в которой происходит связывание и превращение субстрата.
Активный центр образуется радикалами аминокислотных остатков полипептидной цепи при формировании ее третичной структуры; у двухкопонентных ферментов в состав активного центра входят и некоторые группировки небелковой части. Достройка активного центра двухкомпонентных ферментов происходит после взаимодействия апофермента с небелковой частью. Нарушение третичной структуры фермента под влиянием различных факторов приводит к дефомации активного центра и изменению ферментативной активности.
Наиболее часто в состав активных центров ферментов входят радикалы серина, гистидина, треонина, цистеина, аргинина, аспарагиновой и глутаминовой кислот.
Активный центр функционально неоднороден; в нем условно выделяют “каталитически активный” участок, где происходит превра-щение субстрата (расщепление или синтез связи), и так называемый контактный или “якорный” участок, который обеспечивает связывание субстрата с ферментом.
Р ис
4.1 Модель молекулы фермента
ис
4.1 Модель молекулы фермента
А - третичная структура молекулы; Б - силуэт молекулы с активным центром (в рамке).
В молекуле фермента может присутствовать а л л о с т е р и ч е с к и й центр, представляющий собой участок молекулы, присоединение к которому определенных веществ приводит к изменению третичной структуры молекулы фермента. В результате этого происходит изменение конфигурации активного центра, сопровождающееся либо увеличением, либо снижением каталитической активности фермента. Это явление лежит в основе так называемой аллостерической регуляции активности ферментов. Ферменты, активность которых регулируется веществами, присоединяющимися к аллостерическому центру, получили название а л л о с т е р и ч е с к и х ферментов.
4.3 Механизм ферментативного катализа
Химическая реакция имеет определенный “энергетический барьер” и может произойти только в том случае, если реагенты (реагирующие молекулы) обладают запасом энергии, достаточным для достижения ими вершины этого барьера и перехода в промежуточное состояние, называемое а к т и в и р о в а н н ы м комплексом или п е р е х о д н ы м с о с т о я н и е м. В переходном состоянии возможно одновременное образование новых и разрыв старых химических связей.
Активация молекул может происходить при повышении температуры, в результате поглощения ими лучистой энергии, при столкновении с другими возбужденными молекулами или атомами,передающими им часть своей энергии. Количество энергии, необходимое для достижения при данной температуре всеми молекулами одного моля вещества переходного состояния, соответ-ствующего вершине энергетического барьера, называется э н е р г и е й а к т и в а ц и и. Иначе, энергия активации представляет собой “энергетический барьер”, который нужно преодолеть для того, чтобы произошла реакция.
В присутствии катализатора понижается энергия активации . Причем фермент снижает энергию активации значительно сильнее, чем неорганический катализатор.
Согласно теории Михаэлиса-Ментен фермент (Е) соединяется со своим субстратом (S), образуя нестойкий промежуточный комплекс (ES), который в конце реакции распадается с освобождением фермента и продуктов реакции. Поэтому всю последовательность каталитического ферментативного процесса можно представить следующим образом:







 E
+ S ES EX
EP E + P,
E
+ S ES EX
EP E + P,
где ЕХ - истинный активированный комплекс (переходное состояние); ЕР - комплекс фермента с продуктом.
В самой общей форме можно сказать, что молекула субстрата, после связывания с активным центром фермента, поляризуется, электроны в ней перераспределяются, расположение электрических зарядов изменяется, связи деформируются и все это приводит к повышению ее активности.
4.4. Обратимость действия ферментов
Реакции, катализируемые многими ферментами обратимы, т.е. один и тот же фермент в зависимости от определенных условий может катализировать реакцию в обоих направлениях. В 1884г.
Обратимость действия ферментов бесспорно доказана вне организма. Однако в живой клетке большинство ферментативных синтезов (см. обмен веществ) происходит под действием других ферментов, а не тех, которые катализируют расщепление того или иного соединения и, следовательно, не благодаря обратимости действия ферментов. Это, по-видимому, связано с тем, что в живой клетке в большинстве случаев происходит удаление продуктов реакции и, кроме того, реакции синтеза и соответствующие им реакции распада часто локализованы в разных участках, или отсеках (компартментах) клетки. Такая компартментализация гарантирует независимое протекание процессов синтеза и распада и обеспечивает благоприятные энегргетические условия для них.
4.5. Специфичность ферментов
Под специфичностью ферментов понимают способность каждого из них катализировать одну или несколько близких по природе хими-ческих реакций. Это одно из важнейших биологических явлений, без которого невозможен упорядоченный обмен веществ в живом организме, а следовательно, и сама жизнь.
Исследуя природу ферментативного катализа Э.Фишер в 1890-х годах пришел к выводу, что специфичность ферментов можно уподобить соответствию между “ключом и замком”. При этом под-разумевается, что активный центр фермента имеет жесткую структуру, подобно замку. Молекула субстрата должна иметь комплементарную структуру, чтобы входить в активный центр, подобно ключу (рис 4.2). Представление Э.Фишера об активном центре фермента, как жесткой структуре, не подвергалось сомнению в течение полустолетия.
По мере изучения механизма действия ферментов был выявлен ряд данных, которые нельзя согласовать с теорией “ключа и замка”. Например, фермент не может атаковать (подвергнуть превращению) молекулы веществ, обладающие меньшим или большим размером по сравнению с субстратом, но имеющие такие же группы для взаимодействия с ферментом, что и субстрат. Следует отметить, что в ряде случаев такие молекулы связываются с контактным участком активного центра, но при этом превращение субстрата не происходит.
Специфичность у разных ферментов выражена в неодинаковой степени. Различают следующие типы специфичности.
1. А б с о л ю т н а я с п е ц и ф и ч н о с т ь. При этом типе специфичности фермент катализирует превращение только одного субстрата. Фермент каталаза катализирует расщепление пероксида водорода на воду и кислород; ее действие ограничивается только этим субстратом.

Рис 4.2. Схема связывания фермента и субстрата согласно теории “ключа и замка”
Обозначение: E- фермент; S-субстрат; Р - продукты; ES- комплекс фермент-субстрат; ЕХ - истинный активированный комплекс; А - активный центр фермента.
2. Г р у п п о в а я с п е ц и ф и ч н о с т ь. Основным признаком для ферментов этого типа специфичности служит характер разрушаемой или создаваемой связи в близких по строению группах веществ. К ферментам с групповой специфичностью относятся липазы, катализирующие гидролиз сложных эфиров глицерина и карбоновых кислот; фосфатазы, действующие на эфиры фосфорной кислоты; пептидгидролазы, катализирующие гидролиз пептидных связей в белках и пептидах и др.
3. С т е р е о х и м и ч е с к а я с п е ц и ф и ч н о с т ь. Ферменты этого типа специфичности действуют на определенный изомер одного и того же вещества: D- или L-, - или - , транс- или цис- . Пептидгидролазы действуют только на пептиды, образованные аминокислотами L-ряда.
4.6. Кинетика ферментативных реакций
Химическая кинетика - это учение о скоростях и механизмах хи-мических реакций. Ферментативная кинетика изучает закономерности влияния химической природы реагирующих веществ (фермента, субстратов) и условий их взаимодействия (концентрация фермента, концентрация субстратов или ингибиторов) на скорость ферментативных реакций.
4.6.1. Измерение скорости ферментативных реакций
Мерой скорости ферментативной реакции служит количество субстрата, подвергшегося превращению в единицу времени, или количество образовавшегося продукта.
4.6.2. Единицы активности ферментов
Для выражения каталитической активности Комиссией по ферментам Международного биохимического союза (1961 г.) была рекомендована стандартная единица, обозначенная на русском языке - Е, а на английском - U.
С т а н д а р т н а я е д и н и ц а - это такое количество фермента, которое при заданных условиях катализирует превращение одного микромоля субстрата за одну минуту.
В 1972 г. Комиссия по ферментам Международного биохимического союза предложила выражать активность ферментов в к а т а л а х. Катал (символ - кат - это такое количество фермента, которое способно превращать один моль субстрата за одну секунду (при оптимальных условиях).
К производным величинам, характеризующим активность ферментов, относят удельную каталитическую активность ферментов, концентрацию фермента в растворе и другие. Удельную каталитическую активность фермента или ферментативного препарата выражают в каталах на 1 кг белка (кат·кг-1) или чаще в мккат на 1мг белка. Концентрацию фермента в растворе выражают в каталах на 1 литр (кат·л-1) или в других, кратных этому значению величинах.
4.6.3. Влияние концентрации субстрата на скорость ферментативной реакции
Ферментативные реакции, в отличие от неферментативных, обладают очень важной особенностью - насыщения фермента субстратом. Эта особенность проявляется в том, что при постоянной концентрации фермента скорость реакции имеет характерную зависимость от концентрации субстрата. При низких концентрациях субстрата скорость реакциипропорциональна концентрации субстрата и по отношению к нему - это реакция первого порядка. С увеличением концентрации субстрата приращение скорости с каждым разом уменьшается и, наконец, она становится практически независимой от концентрации субстрата. В этих условиях реакция по отношению к субстрату - нулевого порядка, а весь фермент полностью насыщен субстратом и не может функционировать быстрее. Скорость ферментативной реакции при полном насыщении фермента субстратом называется м а к с и м а л ь н о й с к о р о с т ь ю.
Vmax · [S ]
V =
Кm + [S ]
Это окончательное уравнение, выведенное для односубстратной реакции, называют у р а в н е н и е м М и х а э л и с а - М е н т е н. Данное уравнение позволяет легко измерять максимальную скорость из экспериментальных данных, полученных при любой фиксированной концентрации фермента. График зависимости скорости ферментативной реакции от концентрации субстрата представляет собой гиперболу (рис. 4.3).

VmaxРис. 4.3. Зависимость

 скорости реакции
скорости реакции
катализируемой
ферментом, от
1/2 Vmax концентрации
субстрата.
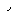
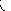



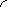 [S] К m
- константа Михаэлиса
[S] К m
- константа Михаэлиса
Km
4.6.4 Влияние концентрации фермента на скорость ферментативной реакции
При высокой концентрации субстрата и при постоянстве других факторов скорость ферментативной реакции зависит от концентрации фермента. При построении графика эта зависимость будет линейной (рис. 4.4).
Рис. 4. Влияние концентрации фермента на скорость реакции: V - скорость реакции; [E] - концентрация фермента.
|
V
E
|
В клетках организма и в производственных условиях катализ всегда осуществляется в условиях, когда концентрация фермента гораздо ниже концентрации субстрата.
|



4.6.5 Влияние температуры на скорость ферментативной реакции
Важным фактором, от которого зависит скорость ферментативной реакции (равно каталитическая активность фермента) является температура, влияние которой показано на рис 4.5. Из рисунка видно, что с повышением температуры до определенной величины скорость реакции увеличивается. Это можно объяснить тем, что с повышением температуры движение молекул ускоряется и у молекул реагирующих веществ оказывается больше возможности столкнуться друг с другом. Это увеличивает вероятность того, что реакция между ними произойдет. Температура, обеспечивающая наибольшую скорость реакции, называется о п т и м а л ь н о й температурой.
Каждый фермент имеет свою оптимальную температуру. В общем для ферментов животного происхождения она лежит между 37 и 40ОС, а растительного - между 40 и 50ОС. Однако есть и исключения: -амилаза из проросшего зерна имеет оптимальную температуру при 60ОС, а каталаза - в пределах 0 - 10ОС. При повышении температуры сверх оптимальной скорость ферментативной реакции снижается, хотя частота столкновений молекул увеличивается. Происходит это вследствие денатурации, т.е. потери ферментом нативного состояния. При температуре выше 80ОС большинство ферментов полностью теряют свою каталитическую активность.
Снижение скорости ферментативной реакции при температурах, превышающих оптимальную, зависит от денатурации фермента. Поэтому важным показателем, характеризующим отношение фермента к температуре, является его термолабильность, т.е. скорость инактивации самого фермента при повышении температуры.
Р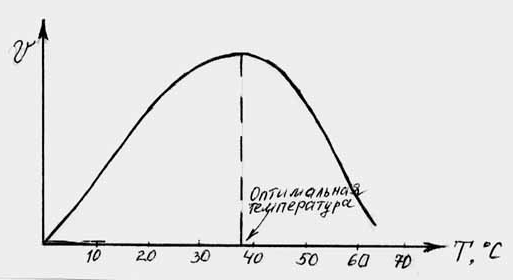 ис.
4.5. Влияние температуры на скорость
гидролиза крахмала амилазой.
ис.
4.5. Влияние температуры на скорость
гидролиза крахмала амилазой.
При низких температурах (0 ОС и ниже) каталитическая активность ферментов падает почти до нуля, но денатурация при этом не происходит. С повышением температуры их каталитическая активность вновь восстанавливается.
4.6.6 Влияние рН на скорость ферментативной реакции
Важным фактором, оказывающим большое влияние на скорость ферментативной реакции, является рН среды.
Для каждого фермента существует оптимальное значение рН, т.е. такая величина рН, или зона рН, при которой катализируемая ферментом реакция протекает с наибольшей скоростью (рис. 4.6).
Большинство ферментов имеют максимальную каталитическую активность в зоне рН от 7; в резко кислой или резко щелочной среде
Рис. 4.6. Влияние рН на
скорость реакции, катализируемой
пепсином (1) и сахарозой из дрожжей (2).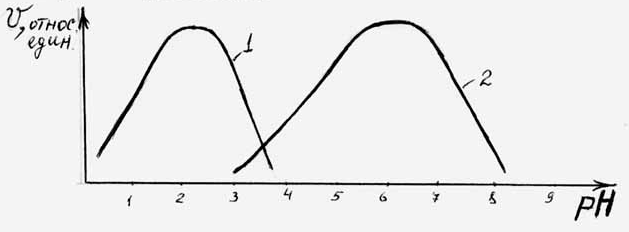
работают лишь некоторые ферменты. За пределами оптимальной зоны рН, т.е. при отклонениях в сторону снижения или в сторону повышения от этого значения, скорость ферментативной реакции снижается.
При разных значениях рН активный центр может находиться в разной степени ионизированной или неионизированной форме, что сказывается на формировании активного фермент-субстратного комплекса. Кроме того, имеет значение факт ионизации субстатов и кофакторов.
При проведении производственных процессов можно путем соблюдения требуемого рН снизить активность нежелательных для процесса ферментов и повысить активность полезных ферментов.
4.6.7 Влияние на каталитическую активность ферментов ингибиторов и активаторов
Скорость ферментативной реакции (равно активность ферментов) определяется присутствием в среде ингибиторов и активаторов , среди которых могут быть как посторонние для организма вещества, так и природные продукты обмена.
И н г и б и т о р а м и называют вещества, вызывающие частичное или полное торможение химических реакций, включая и ферментативные.
Ферменты теряют каталитическую активность при воздействии различных факторов, вызывающих денатурацию (нагревание, кислоты, щелочи, соли тяжелых металлов и др.). Подобное разрушение фермента не рассматривается как ингибирование, так как оно не связано с механизмом действия фермента. Ингибиторы действуют на скорость реакции определенным химическим путем.
Механизм действия ингибиторов может быть самым разнообразным, но в общей форме можно сказать, что ингибитор вступает в соединение с ферментом, образуя соединение фермент-ингибитор.
Различают о б р а т и м о е и н е о б р а т и м о е ингибирование фермента. При обратимом ингибировании активность фермента восстанавливается по мере удаления свободного ингибитора диализом или иным способом, т.е. при обратимом ингибировании существует равновесие между свободным ингибитором и ферментом. При необратимом ингибировании равновесие между свободным ингибитором и ферментом не устанавливается и активность фермента не удается восстановить диализом. Напротив, если ингибитор присутствует в избытке по сравнению с ферментом, то со временем наступает полное торможение активности фермента.
Обратимое ингибирование ферментативных реакций бывает к о н к у р е н т н ы м и н е к о н к у р е н т н ы м.
Конкурентное ингибирование может быть вызвано веществами, похожими по своей структуре на субстрат. Эти вещества, конкурируя с субстратом, соединяются с активным центром фермента, но не подвергаются ферментативному превращению и новые продукты из них не образуются. В связи с тем, что часть фермента при конкурентном ингибировании расходуется на образование комплеса фермент-ингибитор, скорость ферментативной реакции снижается. Конкурентное ингибирование обратимо, так как при увеличении концентрации субстрата скорость реакции возрастает.
Неконкурентное ингибирование вызывают вещества, не имеющие структурного сходства с субстратом. Причем эти вещества обратимо присоединяются к ферменту не в активном центре, где обычно связывается субстрат, а совсем в другом месте и, следовательно, конкуренция между субстратом и ингибитором отсутствует. Связываясь с ферментом, неконкурентные ингибиторы вызывают изменение пространственной структуры активного центра, и, хотя присоединение субстрата к такому активному центру происходит, тем не менее катализ становится невозможным. Неконкурентные ингибиторы связываются обратимо как со свободным ферментом, так и с фермент-субстратным комплексом, образуя неактивные фермент-ингибитор (ЕJ) и (или) фермент-субстрат-ингибитор (ESJ).
Наряду с инигибиторами существует целый ряд активаторов ферментов. А к т и в а т о р а м и называют вещества, увеличивающие каталитическую активность ферментов. Среди активаторов встречаются самые разнообразные вещества. Особенно часто роль активаторов ферментов выполняют ионы металлов: калия, кальция, магния, цинка, меди, железа, марганца, кобальта, а из анионов - хлора. Для проявления максимальной активности ферментов требуется определнная концентрация ионов-активаторов в среде.
Усиление активности ферментов под действием ионов металлов объясняется тем, что в одних случаях ионы металлов выполняют роль кофактора, в других - облегчают образование фермент-субстратного комплекса, в третьих - способствуют прсоединеию кофермента к апоферменту, в четвертых обеспечивают становление четвертичной структуры фермента или же действуют иными путями.
Мощное действие на ферменты оказывают вещества, присоединяющиеся к ним в особых участках, удаленных от активного центра, называемых а л л о с т е р и ч е с к и м ц е н т р о м. Эти вещества влияют на активность фермента, вызывая обратимое изменение в структуре его активного центра. Называют такие вещества а л л о с т е р и ч е с к и м и э ф ф е к т о р а м и. Если эти эффекторы увеличивают сродство фермента к субстрату, то их называют а л л о с т е р и ч е с к и м и активаторами, если уменьшают - а л л о с т е р и ч е с к и м и ингибиторами. Ферменты, активность которых регулируется аллостерическими активаторами или ингибиторами называют а л л о с т е р и ч е с к и м и. Большинство аллостерических ферментов представляют собой белки-олигомеры.
Аллостерические ферменты имеют важное значение в регуляции ферментативных процессов в клетке. Это связано с тем, что эффекторами могут быть различные промежуточные продукты обмена веществ, называемые м е т а б о л и т а м и. В частности, установлено, что конечный, а иногда и промежуточный продукт многостадийного процесса распада или биосинтеза может служить аллостерическим ингибитором одной из первых его реакций.
4.7 Номенклатура и классификаия ферментов
В настоящее время известно более 2400 ферментов. Каждый фермент, как правило, имеет две номенклатуры; одна из них рабочая (тривиальная), а другая - систематическая.
Рабочее наименоваие фермента составляют путем прибавления к корню слова латинского , греческого или химического названия субстрата, на который действует фермент, или к названию процесса, катализируемого данным ферментоа окончания “-аза”. Вещество, имеющее это окончание, принимают за фермент. Ферменты, действующие на крахмал (amylum), сахарозу, мочевину (urea), пептиды получили соответственно названия : амилаза, сахараза, уреаза,пептидаза; ферменты, катализирующие процессы гидролиза называют гидролазами, процессы окисления - оксидазами, перенос групп - трнсферазами и т.д. Для некоторых ферментов сохранены названия, неподчиняющиеся этому правилу: пепсин, трипсин, химотрипсин папин и др.
В названии ряда ферментов указывают как характер субстрата, так и тип катализируемой реакции. Фермент, катализирующий отнятие водорода от спирта, называют алкогольдегирогеназа.
Рабочим названием ферментов пользуются в повседневной практике.
В 1961 г. Международная комиссия по ферментам, созданная в 1956 г., предложила новую схему номенклатуры и классификации ферментов, которая была принята Международным биохимическим союзом. Согласно этой схемы каждый фермент имеет как рекомендуемое (рабочее) название, так и систематическое название, которое составляется в определенном порядке и подчеркивает тип катализируемой реакции (см. классы ферментов).
В принятой классификации все ферменты на основании катализируемых реакций разделены на шесть классов, расположенных в следующем порядке: 1) оксидоредуктазы, 2) трансферазы, 3) гидролазы, 4) лиазы, 5) изомеразы, 6) лигазы (синтетазы). Каждый класс подразделяется на подклассы, а каждый подкласс - на подподклассы. Индивидуальный фермент имеет кодовое число (шифр) со стоящими перед ним буквами КФ (англ. ЕС). Шифр каждого фермента содержит четыре числа, разделенных точками. Первое число указывает к какому из шести классов принадлежит данный фермент. Второе число обозначает подкласс. Третье число обозначает подподкласс, а четвертое - порядковый номер фермента в данном подподклассе. Например, фермент КФ.1.1.1.1 имеет рекомендуемое (рабочее) название алкогольдегидрогеназа, систематическое название алкоголь:НАД оксидоредуктаза. Этот фермент относится к классу оксидоредуктаз (1), действует на СНОН группу доноров (1.1), акцептором водорода служит НАД (1.1.1); четвертая цифра шифра - порядковый номер фермента в пределах подподкласса.
Систематическое название и шифр фермента используют в научных публикациях при первом упоминании о нем; при дальнейшем изложении материала пользуются рекомендуемым (рабочим) названием.
4.8 Классы ферментов и их отдельные представители
4.8.1.Оксидоредуктазы(1)
К классу оксидоредуктаз принадлежат все ферменты, катализирующие окислительно-восстановительные реакции. Субстрат, подвергающийся окислению рассматривается как донор водорода. Систематическое назва-ние составляется по типу “донор:акцептор оксидоредуктаза”. Термины “донор” и “акцептор” введены потому, что происходит реакция переноса двух восстановительных эквивалентов в той или иной форме (атомов водо-рода, электронов, гидрид-ионов и т.д.) от одного субстрата (окисляемого) к другому (восстанавливаемому).
В зависимости от природы окисляемых групп в молекуле донора оксидоредуктазы разделили на 19 подклассов. Деление на подподклассы произведено в зависимости от природы акцепторов, которыми могут быть кофермент (НАД или НАДФ), цитохром, молекулярный кислород и т.д.
Рекомендуемое название включает следующие термины. Оксидоредуктазы всех подподклассов, для которых акцептором водорода служит любое соединение, но не кислород, называют дегидрогеназами, а в случаях, когда донор водорода точно не установлен, используют термин редуктаза. Если акцептором служит кислород, то ферменты, катализирующие перенос водорода на него, называют оксидазами. Реакции прямого включения кислорода в молекулу органического субстрата катализируют оксигеназы; при этом происходит включение либо двух атомов кислорода (диоксигеназы), либо одного атома кислорода (монооксигеназы). Термин пероксидаза относится к ферментам, использующим в качестве окислителя пероксид водорода.
Многие дегидрогеназы в качестве акцептора водорода используют коферменты НАД (никотинамидадениндинуклеотид) и НАДФ (никотинамидадениндинуклеотидфосфат), содержащие в своих молекулах производ-ное пиридина - никотинамид. В связи с чем эти ферменты получили название пиридиновых (пиридинзависимых) дегидрогеназ, или пиридин-протеинов. Приводим структурную формулу окисленной формы НАД и НАДФ (обозн. НАД+ и НАДФ+ ).
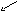
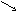 OНикотинамид
АденинNH2
OНикотинамид
АденинNH2
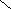

 CNH2
N N
CNH2
N N

 O
O
O
O



 +
N
+
N







 N
O H2CO
P
O
P
O
CH2
O N
N
O H2CO
P
O
P
O
CH2
O N



 H H H
H ОН ОН H H H
H
H H H
H ОН ОН H H H
H
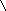
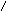
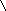
OH OHOH OR
R = Н (в НАД+) и РО(ОН2) (в НАДФ+).
Пиридиновые дегидрогеназы отнимают от своих субстратов по два водородных атома. Один из них в виде гидрид-иона (Н) присоединяется непосредственно к пиридиновому кольцу НАД+ или НАДФ+ , а второй в виде Н+-иона переходит в среду:

 O
H H O
O
H H O

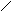


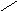 СН
C
СН
C
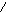



 Н
CNH2
C NH2
Н
CNH2
C NH2
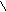
 Субстрат
+ Субстрат
+ +H+
Субстрат
+ Субстрат
+ +H+
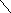


 Н
+ (окисленный)
Н
+ (окисленный)
N N
RR
(Восстановленная форма нуклеотидных коферментов обозначается НАД·Н + Н+ (сокр. НАД·Н) и НАДФ·Н+Н+ (сокр. НАДФ·Н).
В общем виде реакции, катализируемые пиридиновыми дегидро-геназами, можно записать следующим образом:
 Н
НАД+НАД·Н+Н+
Н
НАД+НАД·Н+Н+
С убстрат
+
Субстрат +
убстрат
+
Субстрат +
 Н
(НАДФ+)
(окисленный) (НАДФ·Н+Н+)
Н
(НАДФ+)
(окисленный) (НАДФ·Н+Н+)
При каталитическом участии фермента трансгидрогеназы НАД·Н и НАДФ+, равно как НАДФ·Н и НАД+ , могут обмениваться атомами водорода:
Н
 АДФ·Н
+ НАД+
НАДФ+
+ НАД·Н
АДФ·Н
+ НАД+
НАДФ+
+ НАД·Н
Установлено, что большая часть пиридиновых дегидрогеназ функционирует только с коферментом НАД, меньшая - только с НАДФ и сравнительно небольшая группа - как с НАД, так и с НАДФ.
Считается, что восстановительные эквиваленты от НАД·Н расходуются для запаса энергии в виде АТФ, а от НАДФ·Н для восстановительных этапов процессов биосинтеза.
Пиридиновые дегидрогеназы называют анаэробными, т.к. они передают отнятый от субстрата водород любому соединению, но не кислороду.
Наряду с пиридиновыми дегидрогеназами в окислительно-восстановительных реакциях участвуют флавиновые (флавинзависимые) дегидрогеназы, или флавопротеины. Такое название эти ферменты получили в связи с тем, что в качестве простетической группы (прочносвязанного с белковой частью кофермента) содержат флавинмононуклеотид (ФМН) или флавинадениндинуклеотид (ФАД):
O О
NH2

 СН2
(СНОН)СН2
ОР
О
РОН
СН2
(СНОН)СН2
ОР
О
РОН
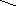



N
N
N
N



 N
N OH О
N
N OH О







 H3C
=O
N
H3C
=O
N








 NH
CH2
O N
NH
CH2
O N
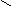 H3C
N
H3C
N



 H
H H H
H
H H H
O
OH
OН 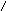
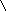



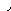


 Рибофлавин
Рибофлавин







 ФМН
(окисленный)
АМФ
ФМН
(окисленный)
АМФ

 ФАД
(окисленный)
ФАД
(окисленный)
Большинство флавиновых дегидрогеназ содержит в своем составе ФАД.
Катализ окислительно-восстановительных реакций флавиновыми дегидрогеназами обусловлен последовательным окислением и восстановлением изоаллоксазинового кольца рибофлавина:
R R H





 N
N N
N
N
N N
N
H







 3C
3C



 =
O +
2H H3C
=
O +
2H H3C



 =O
=O





H






 3C
3C

 NH H3C
NH
NH H3C
NH

 N
C N
C
N
C N
C
O H O
Окисленная форма Восстановленная форма
К а т а л а з а (КФ 1.11.1.6; пероксид водорода:пероксид водорода оксидоредуктаза) катализирует следующую реакцию:
 Н2О2
+ Н2О2
2Н2О
+ О2
Н2О2
+ Н2О2
2Н2О
+ О2
Каталаза - двухкомпонентный фермент, простетической группой которого является гематин, представляющий протопорфирин, содержащий атом трехвалентного железа (см. формулу на стр. 134).
Этот фермент широко распространен в природе и найден у животных, растений, аэробных бактерий. Роль каталазы в организме связывают с расщеплением образующегося в процессе окисления ядовитого для клеток пероксида водорода.
Каталаза содержится и в молоке. В молоко она переходит из клеток молочной железы, а также вырабатывается содержащимися в молоке бактериями и лейкоцитами.
аталазное число молока от здоровых коров составляет 0,7 - 2,5. Молоко коров с больным выменем и молозиво имеют каталазное число достигающее 15 и выше.
Следующий фермент этого подкласса п е р о к с и д а з а (КФ 1.11.1.7; донор:пероксид водорода оксидоредуктаза). Пероксидаза содержится в тканях животных и растений, молоке, лейкоцитах, некоторых бактериях; катализирует окисление фенолов, аминов, некоторых гетероциклических соединений (например, индола) по схеме:
д онор
+ Н2О2
окисленный донор + 2 Н2О.
онор
+ Н2О2
окисленный донор + 2 Н2О.
Пероксидаза - двухкомпонентный фермент, ее простетическая группа представлена гематином. Гематин каталазы и пероксидазы имеет одинаковое строение. Следовательно, различия в каталитической функции этих ферментов определяются исключительно белковой частью.
Пероксидаза играет важную роль в дыхании растений. В молочной промышленности реакцию на пероксидазу используют для контроля эффективности пастеризации молока (пероксидаза молока инактивируется при температуре около 80ОС). Реакцию на пероксидазу применяют для оценки свежести мяса птицы (кроме водоплавающей). Свежее мясо дает положительную реакцию; мясо подозрительной свежести - отрицательную.
Ц и т о х р о м н а я с и с т е м а. Эта система состоит из цитохромов и фермента цитохромоксидазы. Цитохромы принадлежат к сложным белкам; их железопорфириновая простетическая группа, называемая гем, по своему строению очень близка к простетической группе гемоглобина (см. хромопротеины).
Все известные цитохромы в зависимости от природы гема разделены на четыре группы: а, в, с и d. Железопорфириновые структуры каждого из этих цитохромов различаются боковыми цепями. Кроме этого цитохромы отличаются друг от друга белковыми компонентами и по способу присоединения простетической группы к белку. У цитохромов с хорошо установленной структурой при букве ставят числовой индекс, указывающий на принадлежность цитохрома к определенной подгруппе. Например, в митохондриях высших животных и растений идентифицировано пять различных цитохромов: а, а3, в, с, с1.
Цитохромы - переносчики электронов в процессах окисления и восстановления. Они обнаружены во всех аэробных клетках. В ходе переноса электронов железо простетической группы цитохромов попеременно переходит из ферриформы [Fе (III)] в ферроформу [Fе (II)]. Функция цитохромов была установлена в 1925 г. Д.Кейлином.
Цитохромоксидаза, классифицируемая и как фермент (КФ 1.9.3.1; ферроцитохром С: кислород оксидоредуктаза), и как цитохром ( цитохром а а3), содержит две железопорфириновые группы (гем а и гем а3) и два атома меди. Ее функция состоит в переносе электронов на молекулярный кислород; последний при этом приобретает способность реагировать с находящимися в водной среде клетки ионами водорода, образуя воду.
4.8.2. Т р а н с ф е р а з ы ( 2 )
К классу трансфераз принадлежат ферменты, катализирующие перенос различных остатков или групп от одного соединения, расматриваемого как донор группы (остатка), к другому соединению, рассматриваемому как акцептор. Систематическое название ферментов этого класса формируется по схеме “ донор: акцептор (транспортируемая группа или остаток ) трансфераза”.
В зависимости от характера переносимых остатков (одноуглеродные, альдегидные и кетонные, ацильные, гликозильные и др.) или групп (содержащие азот, фосфор или серу) класс разделили на восемь подклассов. Подклассы выделены в зависимости от химической природы переносимых групп (например, одноуглеродный остаток может быть метилом, формилом или карбоксилом; гликозильный остаток - гексозилом или пентозилом и т.п.).
Среди трансфераз имеются ферменты, катализирующие реакции с элементами синтеза. Чтобы подчеркнуть элемент синтеза в катализируемой реакции, для названия таких ферментов (как и фермент других классов, кроме синтетаз) применяют термин “ синтаза “. Рассмотрим некоторые из таких ферментов.
Г л и к о г е н (крахмал) - с и н т а з а (КФ 2. 4.1.11; УДФглюкоза: гликоген 4--глюкозилтрансфераза) катализирует следующию реакцию:
УДФглюкоза + (1,4--D-глюкозил)n (1.4--D-глюкозил)n+1 + 1 +УДФ
В зависимости от синтезируемого продукта рекомендуемое название фермента уточняется. Биосинтез крахмала катализирует крахмалсинтаза, гликогена - гликогенсинтаза.
В реакциях, катализируемых рядом “синтаз” донором сахара являются нуклеозиддифосфатсахара, например, уридиндифосфатглюкоза (УДФ-глюкоза).
О
CH2OH





 H
O H
H
O H



 NH
O O H
NH
O O H

 H
HO
H
HO
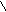
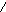




 O=
H2COPO
PO
OH
O=
H2COPO
PO
OH
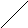


 N O
OH H
N O
OH H
OH OH
H H H H



OH OH Уридиндифосфатглюкоза (УДФ-глюкоза)
4.2.3. Гидролазы (3)
Ферменты этого класса катализируют реакции гидролиза, т.е. расщепление сложных соединений на более простые с присоединением ионов воды: RR1 + HOH = ROH + R1H. В зависимости от типа гидроли-зуемой связи (сложноэфирная, гликозидная, пептидная и т.д.)гидролазы раз-делены на 11 подклассов. Подподклассы выделены с учетом природы суб-страта.
Систематическое название ферментов класса гидролаз составляется из названия гидролизуемого субстрата и названия отщепляемой группы в сочетании с термином “гидролаза”. Гидролазы имеют огромное значение не только для живых организмов, но и для биосферы в целом. Без этих ферментов невозможен круговорот биогенных элементов. Многие из гидролаз имеют промышленное значение.
4.8.4. Лиазы (4)
Ферменты этого класса катализируют удаление из субстратов определенных групп (СО2, Н2О, NH3, альдегид) путем простого отщепления с образованием двойной связи или присоединение группы к двойной связи. Систематическое название фермента складывается из названия субстрата, названия удаляемой группы и через дефис слова “лиаза”. В рекомендуемых названиях используется термин “декарбоксилаза”, “альдолаза”, “дегидратаза”. В зависимости от типа разрываемых связей (С-С, С-О, С-N, C-S и др.) выделено семь подклассов. Деление на подподклассы произведено в зависимости от отщепляемой группы.
4.8.5. Изомеразы (5)
Ферменты этого класса катализируют изомеризацию, т.е. геометрические или структурные изменения, происходящие в пределах одной молекулы. Эти изменения могут происходить вследствие внутримолекулярного перемещения атомов водорода, фосфатных и ацильных групп, различных радикалов, двойных связей и т.п. В зависимости от типа катализируемой реакции изомеразы разделены на пять подклассов; подподклассы выделены в зависимости от характера превращения субстрата.
Фермент, катализирующий внутримолекулярный перенос групп называют мутазой. Изомеразы, катализирующие реакции инверсии при центрах асимметрии, называют рацемазами и эпимеразами. Рацемазы катализируют взаимные превращения D- и L-изомеров, эпимеразы - реакции изменения взаимного расположения атома водорода и гидроксильной группы у одного из углеродных атомов моносахоридов или их производных.
4.8.6. Л и г а з ы , или с и н т е т а з ы ( 6)
Эти ферменты катализируют реакции присоединения друг к другу двух молекул, сопряженные с гидролизом пирофосфатной связи в АТФ или другом нуклеозидтрифосфате. Образуемые в этих реакциях связи часто являются высокоэнергетическими. В зависимости от типа вновь образуемой связи ( - С- О-, -С-S-, -С-N- и др.) лиазы разделили на пять подклассов. Подподклассы выделены в зависимости от природы образующегося соединения. Систематическое название составляется по схеме Х: У лигазы (образующая Z ), где Х и У - соединяющие молекулы; в скобках указывается продукт расщепления нуклеозидтри- фосфата, участвующего в данной реакции в качестве источника энергии. В рекомендуемых названиях применяется термин “синтетаза”.
4.9. Мультиферментные системы.
М у л ь т и ф е р м е н т н ы е с и с т е м ы - это комплексы разных ферментов, катализирующих последовательные стадии превращения какого-либо субстрата. В качестве примера приводим п и р у в а т д е г и д р о г е н а з н у ю с и с т е м у, катализирующую сложный многостадийный процесс - окислительное декарбоксилирование пирувата (см. обмен углеводов), описываемое уравнением:
Пируват + НS-КоА + НАД+ Ацетил- КоА+СО2+ НАД•Н + Н+
Пируватдегидрогеназная система является структурной единицей с молекулярной массой 1·106 - 9·106 в зависимости от биологического ис- точника, состоящей из множества копий (молекул) трех разных ферментов: пируватдегидрогеназы (КФ 1.2.4.1) липоат-ацетилтрансферазы (КФ 2.3.1.12), липоамид-дегидрогеназы (КФ 1.6.4.3) и пяти кофакторов: тиамин-пирофосфата (ТПФ), флавинадениндинуклеотида (ФАД), кофер-мента А ( НS-КоА), никотинамидадениндинуклеотида (НАД+) и
липоевой кислоты ( СН2 - СН2 - СН - (СН2)4 СООН).
 S
S
S
S
Каждый из ферментов пируватдегидрогеназной системы катализирует разные стадии многостадийного процесса. В растительных и животных организмах функционируют и другие мультиферментные системы.