

Болотов. Биология размножения и развития, 2011
.pdfV. Гаметогенез |
101 |
Нормальная продолжительность менструального цикла от 21 до 35 дней, продолжительность менструации в среднем 3–4 дня, до 7 дней, величина кровопотери 50–100 мл. Нормальный менструальный цикл всегда овуляторный, отсутствие выброса в брюшную полость ооцита 2-го порядка говорит об ановуляторном цикле. Однако под влиянием эндогенных и экзогенных факторов может наблюдаться ускорение или замедление овуляции, вплоть до исчезновения её. Обычно у большинства женщин наблюдается от 11 до 13 менструаций в год.
На ускорение овуляции влияет повышенная масса тела, употребление пищи с фитогормонами.
На замедление овуляции влияют: болезни, стресс, физические нагрузки, недостаточное питание, употребление анаболических стероидов, повышенного уровня андрогенов экзогенного или эндогенного происхождения.
Ановуляция может возникнуть по следующим причинам: избыточное содержание в женском организме мужских половых гормонов; заболевания щитовидной железы (гипотиреоз, гипертиреоз); избыточная масса тела; значительное снижение массы тела; чрезмерные физические нагрузки; употребление анаболических стероидов; курение; лекарства: антибиотики, гормоны; алкоголь (регулярное употребление); употребление избытка сладкого; недостаток веса женщины – 45 кг и ниже; опухоли гипофиза и гипоталамуса.
Рис. 20. Регуляция половой функции у женщин
Природой задумано так, что длина первой фазы цикла может очень сильно колебаться у одной и той же женщины, в то же время длина второй фазы остается стабильной, т. е. если овуляция происходит всегда за 15 дней до начала нового цикла
(рис. 20; 21).
Согласно современным представлениям менструальная функция регулируется нейрогуморальным путем
102 |
Болотов А. В. Биология размножения и развития |
при участии: 1) коры головного мозга; 2) подкорковых вегетативных центров, расположенных главным образом в гипоталамусе; 3) гипофиза; 4) яичников; 5) матки, маточных труб и влагалища, содержащих рецепторы, реагирующие на действие половых гормонов яичников. Циклические функциональные изменения в системе гипоталамус – гипофиз – яичники условно объединяются в яичниковый цикл, а циклические изменения в слизистой оболочке матки – в маточный. Одновременно происходят циклические сдвиги во всем организме женщины (менструальная волна), представляющие собой периодические изменения деятельности ЦНС, обменных процессов, функции сердечнососудистой системы и терморегуляции (рис. 21).
Кора головного мозга регулирует процессы, связанные с развитием менструальной функции. Через нее осуществляется влияние внешней среды на нижележащие отделы нервной системы, участвующие в регуляции менструального цикла.
Вгипоталамусе концентрируется влияние импульсов нервной системы и гормонов периферических желез внутренней секреции. В его клетках содержатся рецепторы для всех периферических гормонов, в т. ч. эстрогенов и прогестерона. Нейрогормоны гипоталамуса, стимулирующие освобождение тропных гормонов в передней доле гипофиза, называют рилизингфакторами или либеринами. Существуют также статины – нейрогормоны гипоталамуса, ингибирующие освобождение тропных гормонов в аденогипофизе. Секреция Рилизинг-гормон ЛГ (РГ-ЛГ люлиберин) генетически запрограммирована и происходит в определенном пульсирующем ритме с частотой примерно один раз в час (цирхоральный ритм).
Впередней доле гипофиза (аденогипофиз) синтезируется гонадотропные гормоны гипофиза: фолликулостимулирующий (ФСГ), лютеинизирующий (ЛГ), пролактин (лактотропный, ПРЛ). ФСГ вызывает развитие и созревание первичного фолликула. ФСГ стимулирует рост фолликула, пролиферацию гранулезных клеток, индуцирует образование рецепторов ЛГ на поверхности клеток гранулезы. ЛГ стимулирует образование андрогенов (предшественников эстрогенов) в тека-клетках, совместно с ФСГ способствует овуляции и стимулирует синтез прогестерона в лютеинизированных клетках гранулезы овулировавшего фолликула.

V. Гаметогенез |
103 |
Рис. 21. Гормональная регуляция половой функции у женщин, температурная кривая.
ОВЦ – овуляторный цикл; АВЦ – ановуляторный цикл
Пролактин стимулирует синтез и секрецию прогестерона, превращает не функционирующее желтое тело в функционирующее. При отсутствии пролактина наступает обратное развитие этой железы. Пролактин оказывает многообразное действие на организм женщины. Его основная биологическая роль – рост молочных желез и регуляция лактации. Он обладает также жиромобилизующим эффектом и оказывает гипотензивное действие. Увеличение секреции пролактина является одной из частых причин бесплодия, так как повышение его уровня в крови тормозит стероидогенез в яичниках и развитие фолликулов.
104 |
Болотов А. В. Биология размножения и развития |
В период полового созревания организма фолликулярные клетки зернистого слоя вырабатывают половые гормоны (фолликулины или эстрогены), которые скапливаются в небольших щелях между клетками зернистого слоя. Такой фолликул, в котором имеются полости с гормонами, называется третичным или пузырчатым и наиболее крупные третичные фолликулы с большой полостью внутри и значительным количеством фолликулярных клеток называются Граафовым пузырём. Разрыв зрелого фолликула (овуляция) происходит под влиянием ФСГ, который, влияя на стенку зрелого фолликула, побуждает к выработке в нем гармона эстрина и ЛГ. Происходит разрыв фолликула и выброс ооцита 2-го порядка в брюшную полость. Ооцит 2-го порядка, окруженный фолликулярным эпителием, из брюшной полости попадает в воронку и далее в просвет маточной трубы. Здесь (при наличии мужских половых клеток) быстро происходит второе деление созревания и образуется зрелая яйцеклетка, готовая к оплодотворению.
Значительное число фолликулов не достигает стадии зрелости, а претерпевает атрезию – своеобразную перестройку деструктивного характера. Атрезия ооцитов и возникает атретическое тело (corpus atreticum), внешне несколько напоминающее желтое тело, но отличающееся от последнего наличием в центре блестящей зоны ооцита. В ходе дальнейшей инволюции атретических тел на их месте остаются скопления интерстициальных клеток. Так как интерстициальные клетки участвуют в выработке овариальных гормонов, следует допустить, что атрезия, результатом которой является увеличение количества этих клеток, необходима для гормонообразования в яичнике женщины.
Таким образом, в регуляции функции репродуктивной системы основными являются цирхоральная секреция РГ-ЛГ в нейронах гипоталамуса и регуляция выделения ЛГ и ФСГ эстрадиолом по механизму отрицательной и положительной обратной связи.
Признаки овуляции делятся на объективные и субъективные
Объективные признаки овуляции (эти признаки основыва-
ются на лабораторных исследованиях. При помощи объективных признаков можно точно определить наступление овуляции)
и субъективные признаки овуляции:
V. Гаметогенез |
105 |
1.Общая реакция организма на разрыв фолликула.
2.Гормональный изменения, сопровождающиеся межменструальными болями.
3.Изменение слизи, выделяемой шейкой матки.
4.Изменение шейки матки.
5.Уменьшение времени редукции оксигемоглобина.
6.Увеличение α и β-глобулинов.
7.Уменьшение альбуминов.
8.Повышение секреции гонадотропных гормонов.
9.Межменструальные кровотечения.
10.Падение экскрекции витамина С.
11.Увеличение либидо (половое влечение).
12.Болезненность груди.
13.Боль внизу живота.
14.Изменение шейки матки и ее выделения.
15.Изменение настроения (депрессия).
16.Изменение чувствительности рецепторных элементов. Как правило, чувствительность повышается.
17.Повышение ректальной Т ºС в фазе желтого тела и ↓Т ºС перед овуляцией: Обычно ректальная Т ºС < 37 ºС, за 4–6 дней до овуляции начинает расти, затем за 1–3 дня до овуляции ↓Т ºС. После овуляции (фаза желтого тела) Т ºС q на 0,5–0,8 ºС и держится в течение всей второй фазы, снижаясь лишь за 1–2 дня до менструации (рис. 21).
Двухфазная температурная кривая указывает на чередование фолликулиновой и прогестероновой фаз и говорит об овуляторном цикле. В случае однофазной температурной кривой можно говорить об ановуляторном цикле.
Желтое тело (corpus luteum)
Под воздействием ЛГ образуется желтое тело (рис. 9, 8–9, 11) и синтезируется прогестерон в лютеинизированных клетках гранулезы овулировавшего фолликула.
Под влиянием избытка лютеинизирующего гормона, вызвавшего овуляцию, элементы стенки лопнувшего зрелого пузырька претерпевают изменения, приводящие к формированию желтого тела – временной добавочной эндокринной железы в составе яичника. При этом в полость запустевшего пузырька изливается кровь из сосудов внутренней оболочки, целость кото-
106 |
Болотов А. В. Биология размножения и развития |
рых нарушается в момент овуляции. Сгусток крови быстро замещается соединительной тканью в центре развивающегося желтого тела.
Вразвитии желтого тела различают четыре стадии: 1) пролиферации; 2) железистого метаморфоза; 3) расцвета; 4) инволюции.
Впервой стадии – пролиферации и васкуляризации – происходит размножение эпителиоцитов бывшего зернистого слоя и между ними интенсивно врастают капилляры из внутренней оболочки. Затем наступает вторая стадия – железистого метаморфоза, когда клетки фолликулярного эпителия сильно гипертрофируются и в них накапливается желтый пигмент (лютеин), принадлежащий к группе липохромов. Такие клетки называются лютеиновыми или лютеоцитами (luteocyti). Объем новообразующегося тела быстро увеличивается, и оно приобретает желтый цвет. С этого момента желтое тело начинает продуцировать свой гормон – прогестерон, переходя, таким образом, в третью стадию – расцвета. Продолжительность этой стадии различна. Если оплодотворения не произошло, период расцвета желтого тела ограничивается 12–14 днями. В этом случае оно называется менструальным желтым телом (corpus luteum menstruationis). Бо-
лее длительно желтое тело сохраняется, если наступила беременность – это желтое тело беременности (corpus luteum graviditationis).
Разница между желтым телом беременности и менструальным ограничивается только длительностью периода расцвета и размерами (1,5–2 см в диаметре у менструального и более 5 см в диаметре у желтого тела беременности). После прекращения функционирования как желтое тело беременности, так и менструальное претерпевают инволюцию (стадию обратного развития). Железистые клетки атрофируются, а соединительная ткань центрального рубца разрастается. В результате на месте бывшего желтого тела формируется белое тело (corpus albicans) – соединительнотканный рубец. Оно сохраняется в яичнике на протяжении нескольких лет, но затем рассасывается.
V. Гаметогенез |
107 |
Под влиянием прогестерона происходит увеличение эндометрия (слизистой матки) с 1 мм до 7 мм. Он становится отечным, маточная железа приобретает штопорообразный вид. Просвет переполнен секреторными продуктами. Такие изменения необходимы для имплантации зародыша в стенку матки. В маточных трубах и влагалище имеются также рецепторы к половым стероидным гормонам, но циклические изменения в них выражены менее отчетливо. В других органах (молочные железы, кожные покровы, кости, жировая ткань) клетки названных тканей и органов содержат рецепторы к половым гормонам. В предменструальном периоде количество железистых ходов в молочной железе увеличивается, они расширяются, доли становятся отечными, эпителиальный слой набухает, вакуолизируется и может ощущаться болезненность. В послеменструальном периоде отечность долей, инфильтрация вокруг крупных ходов исчезают и неприятные ощущения тоже исчезают.
В головном мозге также обнаружены рецепторы к половым гормонам, что, по-видимому, может объяснять циклические колебания психики женщины в течение менструального цикла.
8. Сперматогенез
Мужские половые клетки, как и женские, возникают из первичных гоноцитов. При сперматогенезе непосредственными потомками гоноцитов являются стволовые сперматогенные клетки (у млекопитающих называемые также сперматогониями типа А) (рис. 22, Б). Они присутствуют не только у зародышей, но и у половозрелых самцов; в семенниках млекопитающих они располагаются в пристеночном слое семенных канальцев. Стволовые клетки время от времени (нерегулярно) делятся, оставаясь в недифференцированном состоянии. Некоторые из них при этом перемещаются ближе к центру семенного канальца, их деления становятся более регулярными, а после каждого деления эти клетки изменяют свою величину и форму. Такие клетки называют либо просто сперматогониями, либо сперматогониями типа В, а их деления – сперматогониальными (рис. 22, Б).
Сперматогониальные деления постоянно происходят у половозрелых самцов. Однако число делений отдельного сперма-
108 |
Болотов А. В. Биология размножения и развития |
тогония невелико (от 1 до 14) и строго определенно для каждого вида животного. Например, у паразитического червя Dicyema каждый сперматогоний делится всего один раз, у человека, повидимому, 4 раза, а у рыбки гуппи – 14 раз. Откуда же в таком случае берется столь значительное количество продуцируемых сперматозоидов (до 108 в сутки у человека и кролика, 1010 у хряка и лошади и т. д.)? Очевидно, что в основном за счет делений стволовых клеток. И действительно, подсчитано, что у дрозофилы каждая стволовая сперматогониальная клетка (а всего этих клеток порядка 28) делится 107 раз, т. е. продуцирует 2107 сперматогониев, каждый из которых затем делится еще всего 4 раза.
После определенного числа делений сперматогонии передвигается еще ближе к просвету канальца, вступает в профазу 1- го деления созревания и начинает называться сперматоцитом 1- го порядка (рис. 22, Б). В результате 1-го деления созревания
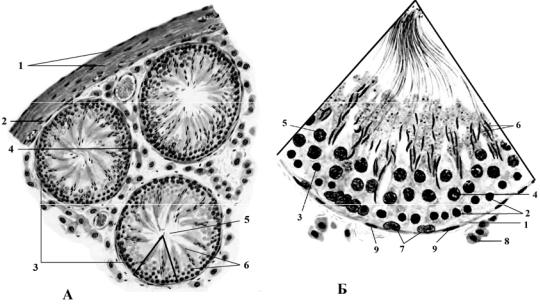
Рис. 22. Семенник, поперечный срез извитых канальцев (А). Часть поперечного среза извитого семенного канальца (Б). Окраска гематоксилин-эозином. х900 (по Алмазову, 1978).
А: 1 – белочная оболочка; 2 – сосудистая оболочка; 3 – извитые семенные канальцы; 4 – интерстиций яичка; 5 – просвет извитого канальца; 6 – сперматозоиды на разной стадии развития.
Б: 1 – волокнистый слой; 2 – сперматогонии А; 3 – сперматогонии В; 4 – сперматоциты 1-го порядка; 5 – сперматоциты 2-го порядка; 6 – сперматозоиды; 7 – фолликулярная клетка (клетка Сертоли); 8 – интерстициальные клетки яичка (клетки Лейдига); 9 – миоидные клетки в составе собственной оболочки канальца
V. Гаметогенез |
109 |
сперматоцит 1-го порядка делится на два одинаковых сперматоцита 2-го порядка (рис. 22, Б), а последние в ходе 2-го деления созревания – на две сперматиды, обладающие, как и зрелая яйцеклетка, гаплоидным числом хромосом и количеством ДНК, соответствующим 1с. Затем каждая сперматида в результате сложных цитологических преобразований, не сопровождающихся клеточными делениями, преобразуется в сперматозоид (рис. 22, Б). Этот процесс спермиогенеза продолжается у разных видов животных несколько дней.
Как сперматогонии, так и сперматоциты и сперматиды всех исследованных видов животных почти непрерывно связаны между собой цитоплазматическими мостиками, образуя синцитий. Этим, по-видимому, объясняется высокая степень синхронности делений сперматогониев и сперматоцитов в данном отделе семенного канальца. Деления эти идут последовательными циклами с периодом в несколько дней, а вдоль семенного канальца наблюдается волна сперматогониальных делений и вхождения в спермиогенез.
Большое значение для сперматогенеза имеют соматические клетки, расположенные в стенках семенных канальцев, – так называемые клетки Сертоли (рис. 22, Б). К ним прикрепляются сперматиды и уже сформированные молодые сперматозоиды. По-видимому, клетки Сертоли снабжают сперматогониальные клетки питательными веществами и гормонами, способствуют высвобождению уже сформированных сперматозоидов в просвет канальцев, а также фагоцитируют неполноценные сперматозоиды.
Основные процессы, протекающие в ходе спермиогенеза (преобразования сперматиды в сперматозоид), таковы (рис. 23):
1)ядро сперматиды сильно уплотняется, хроматин конденсируется и становится синтетически совершенно инертным (рис. 23, А);
2)происходят перемещения органелл клетки; аппарат Гольджи также уплотняется, прижимается к ядру и формирует так называемую акросому, смещается на апикальный конец клетки: центриоли, напротив, смещаются на противоположный полюс ядра и располагаются одна ближе к ядру (проксимальная центриоль), другая дальше (дистальная центриоль) (рис. 23, А, Б, В);
110 |
Болотов А. В. Биология размножения и развития |
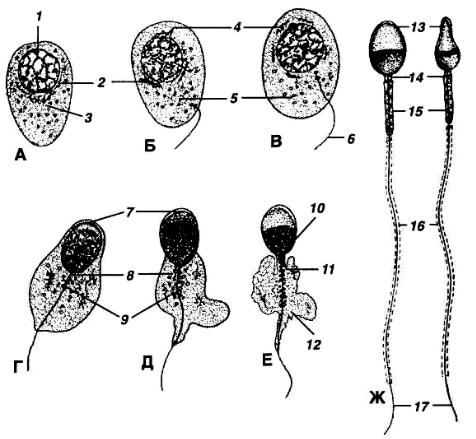
Рис. 23. А – Ж – последовательные фазы спермиогенеза.
1 – ядро сперматиды; 2 – аппарат Гольджи; 3 – центриоли; 4 – зачаток акросомы; 5 – митохондрии; 6 – жгутик; 7 – акросома; 8 – проксимальная центриоль; 9 – дистальная центриоль; 10 – ядерное вещество, сосредоточенное в головке сперматозоида; 11 – митохондриальная спираль; 12 – остатки цитоплазмы; 13 – головка; 14 – шейка; 15 – средняя часть;
16– хвост; 17 – концевой участок хвоста (по Карлсону, 1983)
3)из дистальной центриоли начинает расти жгутик, представляющий собой орган движения сперматозоида. У некоторых видов сперматозоиды лишены жгутика (круглые черви, ракообразные). Вокруг основания жгутика, если он есть, располагаются митохондрии в виде спирального чехлика; по-видимому, они поставляют энергию для движения жгутика (рис. 23, Б, В, Г, Д);
4)вся или почти вся цитоплазма отторгается; зрелый сперматозоид практически ее лишен (рис. 23, Е, Ж).
Для каждого вида животных время, необходимое для превращения сперматогония в зрелый спермий (включая время пребывания в придатке), постоянно, хотя различия между видами существенны. Продолжительность спермиогенеза составляет, в днях: