

Болотов. Биология размножения и развития, 2011
.pdfV. Гаметогенез |
91 |
1.Стадия размножения происходит у самок-эмбрионов в яичниках. При этом зачатковый эпителий, расположенный на поверхности яичника в виде тяжей, которые позже превращаются в стенки флюгеровских мешков. В эти мешки поступают первичные половые клетки – овогонии, которые многократно делятся путём митоза. Мешки постепенно распадаются на первичные или примордиальные фолликулы. Они состоят из половой клетки овогонии, окружённой одним слоем рыхлорасположенных плоских фолликулярных клеток (это бывшие клетки зачаткового эпителия). Стадия размножения заканчивается к моменту рождения самки. При этом около 200 тысяч первичных или спящих фолликулов располагаются возле наружной оболочки яичника. В течение жизни спящие фолликулы поочерёдно переходят в стадию роста.
2.Стадия роста происходит у родившихся самок в яичниках. У человека протекает в функционирующем зрелом яичнике (после полового созревания девочки) и состоит в превращении ооцита 1-го порядка первичного фолликула в ооцит 1-го порядка
взрелом фолликуле. При этом под влиянием гормонов пробуждается половая клетка, которая теперь называется ооцит 1-го порядка. В стадию роста ооцит 1-го порядка растёт и готовится к будущему делению путём мейоза, т. е. стадия роста является интерфазой перед мейозом.
И начинается весьма своеобразный, только этой клетке свойственный период роста. Он связан с поступлением в яйцеклетку питательных веществ извне и с рядом синтетических процессов в самой яйцеклетке. Увеличение массы и объема яйцеклетки может быть колоссальным. Так, ооциты дрозофилы за 3 дня увеличиваются в 90 тыс. раз. У лягушек диаметр молодого ооцита примерно 50 мкм, а зрелого – 2 000 мкм, что соответствует увеличению объема в 64 тыс. раз. Рост ооцита идет сравнительно медленно: только 3-летние лягушки достигают половозрелости. Несравненно быстрее растет яйцо у птиц. Например, у курицы последние 6 дней перед овуляцией объем яйца возрастает в 200 раз. Яйца млекопитающих меньше по размеру; диаметр яйца мыши возрастает от 20 до 70 мкм, что все же соответствует росту объема более чем в 40 раз. Рост яйцеклетки млекопитающих может длиться десятки лет, например, у человека – около 30 лет.
92 |
Болотов А. В. Биология размножения и развития |
При подготовке к 1-му делению созревания ооцит длительное время находится в фазе диакинеза, когда и происходит его рост и вителлогенез. Рост ооцитов всех животных принято разделять на два периода. Первый из них называют периодом малого роста (стадия Винвартера), превителлогенеза, или цитоплазматического роста; второй – периодом большого роста, вителлогенеза, или трофоплазматического роста. Для первого периода характерно относительно малое и притом пропорциональное увеличение массы ядра и цитоплазмы; таким образам, ядерноцитоплазматическое отношение при этом не изменяется. Второй период характеризуется, напротив, резкой интенсификацией роста цитоплазматических компонентов: ядерно-цитоплазма- тическое отношение при этом уменьшается. В течение второго периода в ооците откладывается желток.
В период вителлогенеза в ооците 1-го порядка образуется желток, а также другие питательные вещества – жиры и гликоген. Желток представляет собой высокофосфорилированный кристаллический белок. Он откладывается в виде желточных гранул, одетых пограничной мембраной. По количеству откладываемого в период вителлогенеза желтка яйцеклетки принято делить на полилецитальные, или многожелтковые (например, большинство членистоногих, некоторые рыбы, птицы), мезолецитальные – со средним количеством желтка (амфибии, осетровые рыбы), олиголецитальные – маложелтковые (большинство червей, моллюсков, иглокожих) и алецитальные, т. е. практически безжелтковые (плацентарные формы – млекопитающие, а также и некоторые беспозвоночные, например первичнотрахейные). Количество желтка строго детерминировано генетически и не зависит от условий питания самки.
По способу образования желток принято разделять на экзогенный и эндогенный. Присущий большинству видов животных экзогенный желток строится на основе белка – предшественника вителлогенина, который поступает в ооцит извне. У позвоночных вителлогенин синтезируется в печени матери и транспортируется к содержащему ооцит фолликулу по кровеносным сосудам. Попадая затем в пространство, непосредственно окружающее ооцит (периооцитное пространство), вителлогенин поглощается ооцитом путем пиноцитоза, в составе так называемых
V. Гаметогенез |
93 |
окаймленных мембранных пузырьков. Каждая гранула желтка образуется в результате слияния большого количества (до тысячи) таких пиноцитозных пузырьков. При формировании гранул желтка вителлогенин расщепляется на сильно фосфорилированный белок фосвитин и другой белок – липовителлин, содержащий до 20 % липидов. Эндогенный желток синтезируется из низкомолекулярных предшественников внутри самого ооцита, в его сильно развитом эндоплазматическом ретикулуме. Желточные гранулы возникают из концевых цистерн аппарата Гольджи. Лишь немногие типы яйцеклеток развиваются исключительно за счет эндогенного желтка.
Подготовка к делениям созревания и синтетические процессы в период превителлогенеза. Весь период превителлогенеза проходит на фоне подготовки ооцита 1-го порядка к последующим делениям созревания, или мейоза. Подготовка к 1- му делению созревания начинается вслед за прекращением оогониальных делений с того, что ооцит вступает в S-фазу редукционного деления, т. е. в фазу удвоения ДНК (предшествующую любому клеточному делению). Удвоенное количество ДНК в геноме, достигнутое по завершении этой фазы (4с). Затем наступает профаза 1-го деления мейоза. Профаза мейоза продолжается у ооцитов млекопитающих от несколько дней (у кроликов – до 20 дней) до нескольких лет (у птиц и млекопитающих), однако по достижении фазы диплотены, когда гомологичные хромосомы уже прошли конъюгацию и начали расходиться, наступает стационарная фаза диакинеза. На этой стадии дальнейшее течение мейоза сильно замедляется или же полностью прекращается. Этот блок мейоза продолжается до достижения особью половозрелости, т. е. у ряда млекопитающих и человека много лет. Однако в период диакинеза ядерный материал ооцита не является инертным: у большинства яйцеклеток, обладающих так называемым солитарным, или фолликулярным способами питания, он выполняет роль матрицы для синтеза всех видов РНК – рибосомных, транспортных и информационных, или матричных (соответственно рРНК, тРНК и мРНК). Все эти типы РНК синтезируются преимущественно впрок, т. е. для использования уже оплодотворенной яйцеклеткой.
94 |
Болотов А. В. Биология размножения и развития |
События, происходящие в половой клетке в это время:
1)происходит накопление питательных веществ, как источника энергии;
2)удваивается молекула ДНК;
3)удваивается клеточный центр;
4)синтезируется специальный белок типа тубулина для построения нитей веретена деления;
5)происходит конъюгация и кроссинговер гомологичных хромосом.
Кроме ооцита 1-го порядка в стадию роста начинают расти
ивспомогательные фолликулярные клетки. С этого момента фолликулы получают название вторичных или растущих фолликулов. Постепенно плоские фолликулярные клетки становятся кубическими, а затем и высокими – призматическими. Они формируют вторичную оболочку – лучистый венец. Затем, за счёт веществ, вырабатываемых ооцитом 1-го порядка и клетками лучистого венца, между ними появляется блестящая или прозрачная оболочка, состоящая из гликозаминогликанов (гиалуроновая кислота) и белков. Затем фолликулярные клетки лучистого венца начинают размножаться и формируют снаружи зернистый слой – это будет третья вторичная оболочка, состоящая из множества плотно прилегающих друг у другу фолликулярных клеток (отсутствуют щели между клетками). Снаружи вторичный фолликул покрывается соединительно-тканной оболочкой – текой. Итак: вторичный или растущий фолликул состоит из овоцита 1-го порядка, покрытого плазмалеммой, блестящей оболочкой, лучистым венцом (это первый слой кубических или призматических фолликулярных клеток). В некоторых фолликулах снаружи лучистого венца расположен зернистый слой из плотно прилегающих фолликулярных клеток.
В период полового созревания организма фолликулярные клетки зернистого слоя вырабатывают половые гормоны (фолликулины или эстрогены), которые скапливаются в небольших щелях между клетками зернистого слоя. Такой фолликул, в котором имеются полости с гормонами, называется третичным или пузырчатым. Самые зрелые третичные фолликулы называются Граафовым пузырём.
V. Гаметогенез |
95 |
Наружная оболочка Граафова пузыря – тека, состоящая из двух слоев: наружного – фиброзного и внутреннего – сосудистого, состоящего из ретикулярной ткани, пронизанной множеством капилляров. Здесь содержится много кровеносных сосудов и интерстициальных клеток, дополнительно вырабатывающих половые гормоны. Тека отделяется от эпителиальных фолликулярных клеток зернистого слоя базальной мембраной (это студневидная прослойка). Внутренний слой зернистой оболочки представлен клетками цилиндрической формы с радиальным расположением. Этот слой называется лучистым венцом (рис. 10, 4). Своими отростками фолликулярные клетки передают питательные вещества яйцеклеткам, которые находятся в стадии роста. Среди клеток зернистого слоя появляются щели, или полости, заполненные жидкостью. Таких щелей становится все больше, они объединяются и образуют общую полость, с этого момента фолликул называется пузырчатым или Граафовым пузырьком. Граафов пузырек сильно увеличивается и выпячивает стенку яичника. Внутри фолликула появляется бугорок (яйценосный) (рис. 10, 1) – это скопление фолликулярных клеток, и на его вершине среди зернистых фолликулярных клеток располагается созревающее яйцо, окруженное высокими фолликулярными трофическими клетками, образующими лучистый венец. Стенки Граафова пузырька секретируют женский половой гормон – эстроген, которым насыщается жидкость пузырька. Попадая в кровь, он стимулирует образование лютеотропного гормона в гипофизе. Скопление половых гормонов в Граафовом пузыре приводит к тому, что пузырь лопается, а ооцит 1-го порядка в прозрачной оболочке и лучистом венце овулирует в брюшную полость (рис. 9, 7), где его подхватывает воронка яйцевода. Дальнейшее развитие половой клетки будет происходить в трубке яйцеводе. У свиньи в одном Граафовом пузырьке может созревать несколько яйцеклеток. У овец могут овулировать несколько Граафовых пузырьков.
3. Стадия созревания, или мейоз, состоит из двух быстро следующих друг за другом делений – редукционного и эквационного (уравнительного). При первом этапе (начало) мейоза ооцит 1-го порядка даёт начало двум дочерним клеткам с половинным набором хромосом. Это будет ооцит 2-го порядка и ма-
96 |
Болотов А. В. Биология размножения и развития |
ленькое направительное тельце. И завершается выходом его из яичника в результате овуляции.
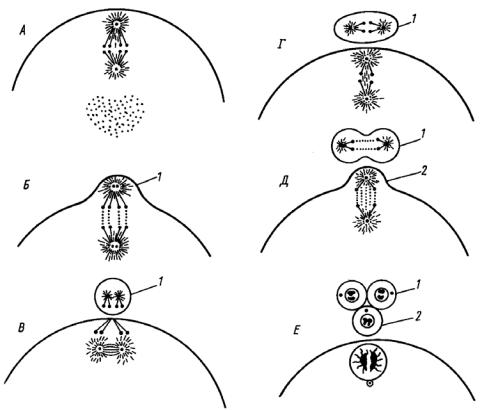
Рис. 18. Схема делений созревания в ооците (по Э. Вильсону, 1940):
А – метафаза 1-го деления созревания; Б, В – выделение I полярного тельца (1); Г, Д – метафаза и анафаза 2-го деления созревания (одновременно делится I полярное тельце); Е – выделение II полярного тельца (2)
Перед 1-м делением созревания ядро ооцита мигрирует к его поверхности. Та точка поверхности ооцита, к которой ближе всего располагается ядро, названа анимальным, противоположная – вегетативным полюсом яйцеклетки. Затем в результате 1-го деления созревания половина хромосомного набора выталкивается в очень маленькую клетку, которая называется первым редукционным, или полярным тельцем (рис. 18, В). Впоследствии эта клетка делится на две столь же малые, и никакого участия в дальнейшем развитии они не принимают (за исключением некоторых случаев партеногенеза). Яйцевая клетка после выделения 1-го редукционного тельца называется ооцитом 2-го порядка. Второе деление созревания осуществляется путем
V. Гаметогенез |
97 |
выделения 2-го редукционного тельца таких же размеров, как и 1-го. После его выделения ооцит 2-го порядка превращается в зрелое яйцо (рис. 18, Г). Биологический смысл резкой неравномерности делений созревания очевиден: невыгодно дробить на части накопленный в процессе роста яйцеклетки запас питательных веществ.
Итак, блок мейоза продолжается до достижения особью половозрелости, Выход из фазы диакинеза и начало собственно делений созревания определяются половыми гормонами: гонадотропные гормоны гипофиза воздействуют на фолликулярный эпителий, продуцирующий в ответ прогестерон или его аналоги. Гормоны фолликулярного эпителия поступают в ооцит и стимулируют его к созреванию.
Современные представления о механизмах этого процесса таковы. Как уже говорилось, в период роста ооцита он связан с фолликулярными клетками щелевыми контактами, через которые из последних в ооцит поступает цАМФ. Высокий уровень цАМФ блокирует (путем фосфорилирования) белковый фактор созревания, который присутствует в цитоплазме ооцита. Под действием прогестерона или его аналогов щелевые контакты размыкаются, концентрация цАМФ в цитоплазме ооцита падает, и фактор созревания деблокируется (дефосфорилируется). Далее он способен еще не вполне изученным образом размножаться (амплифицироваться) в цитоплазме: достаточно инъецировать немного этого фактора в незрелый ооцит, чтобы затем его количество возросло. Фактор созревания вызывает дезинтеграцию (разрушение) оболочки ядра ооцита, разрушение ядрышек и миграцию хромосом к будущему анимальному полюсу, где и произойдут деления созревания.
Из двух делений созревания первое у большинства видов является редукционным, так как именно в ходе этого деления гомологичные хромосомы расходятся по разным клеткам. Таким образом, каждая из разделившихся клеток приобретает половинный (гаплоидный) набор хромосом, где каждый ген представлен лишь одной аллелью. Поскольку 1-му делению созревания предшествовала, как и обычному митотическому делению, S- фаза, каждая из разошедшихся хромосом содержит двойное количество ДНК (две хроматиды). Эти генетически идентичные
98 |
Болотов А. В. Биология размножения и развития |
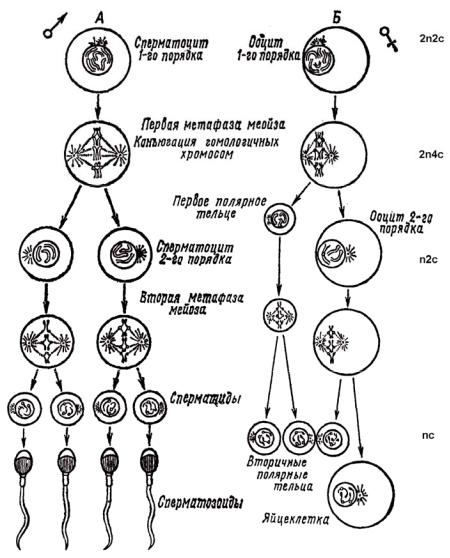
хроматиды и расходятся по сестринским клеткам во 2-м делении созревания, которое является эквационным (как и обычное деление соматических клеток). Наследственный материал представляет из себя одну хромосому, содержащую двойное количество ДНК (1n2c) (рис. 19).
Рис. 19. Схема оогенеза и сперматогенеза
Эти клетки без интерфазы моментально делятся снова, проходя состояние профазы, метафазы, анафазы и телофазы. При этом из ооцита 2-го порядка образуется крупная зрелая яйцеклетка и крошечное направительное тельце. А из первого направительного тельца также образуются две дочерние клетки – два направительных тельца (рис. 18, 2; 19).
V. Гаметогенез |
99 |
Итог мейоза: четыре дочерние клетки с половинным набором хромосом (это яйцеклетка и три направительных тельца, которые почти сразу погибают и рассасываются). На месте лопнувшего Граафова пузыря в яичнике отмечается небольшое кровотечение. Здесь размножаются фолликулярные клетки, откладывается желтый пигмент. Так образуется желтое тело – железа внутренней секреции. Она вырабатывает гормон – прогестерон, тормозящий овогенез. Различают:
1)желтое тело полового цикла, оно функционирует один половой цикл и рассасывается, превращаясь сначала в белое те-
ло (рис. 9, 11);
2)желтое тело беременности (функционирует в период беременности). Некоторые третичные фолликулы в яичнике, наработав гормоны, подвергаются разрушению.
После двух делений созревания число хромосом в каждой из клеток оказывается гаплоидным (1n), количество ДНК в каждом клеточном ядре тоже будет гаплоидным (1с).
Основная особенность делений созревания в ооцитах состоит в том, что эти деления резко неравномерны.
Лишь у немногих видов (некоторые кишечнополостные; из иглокожих – морские ежи) мейоз доходит до конца без участия сперматозоида, внедряющегося в яйцеклетку. У большинства животных течение мейоза останавливается на некотором этапе созревания (возникает, как говорят, блок мейоза), и для дальнейшего его протекания требуется оплодотворение яйцеклетки сперматозоидом (или действие другого активирующего агента).
Различают следующие три типа блока мейоза:
1. Мейоз останавливается на стадии диакинеза, т. е. участие сперматозоида необходимо для протекания обоих мейотических делений. Этот тип блока мейоза наблюдается у губок, отдельных представителей плоских, круглых и кольчатых червей, моллюсков
ищетинкочелюстных. Удивительным образом сюда же относятся три представителя млекопитающих: собака, лиса и лошадь.
2. Мейоз останавливается на метафазе 1-го деления созревания. Такой блок мейоза отмечен у некоторых губок, немертин, кольчатых червей, моллюсков и почти у всех насекомых.
3. Мейоз останавливается на метафазе 2-го деления созревания. Сюда относятся почти все хордовые. У летучих мышей
100 |
Болотов А. В. Биология размножения и развития |
блок мейоза происходит на анафазе 2-го деления созревания. Именно на этих стадиях созревания происходит овуляция яйцеклетки у млекопитающих. Лишь оплодотворение позволяет завершиться ее созреванию и перейти к дальнейшему развитию.
4. Мейоз не блокируется, яйцо останавливается в развитии после завершения мейоза, когда сформирован женский пронуклеус (некоторые кишечнополостные и иглокожие – морские ежи).
В большинстве случаев блок мейоза снимается после активации яйца при оплодотворении или путем искусственной активации. Механизм блока мейоза пока не выяснен.
Яйцеклетки, или ооциты созревают в неизмеримо меньшем количестве, чем сперматозоиды. У женщины в течение полового цикла 24–28 дней) созревает, как правило, одна яйцеклетка. Таким образом, за детородный период образуются около 400 зрелых яйцеклеток.
7.Гормональная регуляция половой функции
уженщин. Овуляция
Следует отметить, что большинство ооцитов в растущих и созревающих фолликулах в разные периоды своего роста претерпевают атрезию. Завершающие этапы периода большого роста ооцита и фолликула, созревания и овуляции происходят циклически и зависят от циклической деятельности системы гипофиз – гипоталамус – яичники.
Женские половые клетки, так же как и мужские, в определенной степени отделены от микроокружения гематофолликулярным барьером, который создает оптимальные условия для метаболизма ооцитов. Он состоит из соединительной ткани (теки), сосудов микроциркуляторного русла, базальной мембраны, фолликулярного эпителия и блестящей зоны.
Вся система регуляции менструального цикла построена по иерархическому принципу (нижележащие структуры регулируются вышележащими, которые, в свою очередь, реагируют на изменения в нижележащих уровнях).
При этом сигналы, поступающие от нижележащих структур, корректируют деятельность вышележащих.