

Петров_К_М_Биогеография_СПб_2001
.pdf(Stipa lessingiana), ковыль-тырса (S. capillata); кроме того типчак (Festuca valesiaca), келерия (Koeleria cristata); полыни (Artemisia).
“Курской растительной аномалией” назвал в начале ХХ века профессор В. В. Алехин степи под Курском. Но эта “аномалийность” связана не с тем, что нетипична растительность, а с тем, что здесь сохранились в условиях сплошной распашки одни из немногих нетронутых целинных участков лу-
говой степи. На этих землях организован Центрально-Черноземный государственный заповедник им. В. В. Алехина.
231

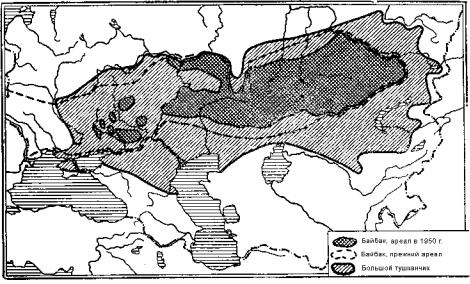
Рис. 9.123. Характерные представители фауны Европейско-Казахских степей. Рис. В. А. Ватагина, по Н. А. Бобринскому:
1 – обыкновенный хомяк; 2 – крапчатый суслик; 3 – степная мышевка; 4 – слепыш; 5 – степная пеструшка; 6 – заяц-русак; 7 – дрофа; 8 – большой тушканчик; 9 – байбак; 10 – степная пищуха; 11 – степной жаворонок; 12 – черный жаворонок; 13 – краснощекий суслик; 14 – стрепет; 15 – сайгак; 16 – полоз желтобрюхий
232

Рис. 9.124. Характерные представители фауны Забайкальских степей. Рис. В. А. Ватагина, по Н. А. Бобринскому:
1 – копытка; 2 – даурская пищуха; 3 – корсак; 4 – манул; 5 – тарбаган; 6 – заяц-толай; 7 – дзэрэн; 8 – даурский суслик; 9 – монгольская песчанка; 10 – даурский цокор; 11 – монольский жаворонок; 12 – монгольская ящурка; 13 – тушканчик-прыгун; 14 – черный журавль
233
Степная растительность заповедника характеризуется богатством видового состава – до 220 видов трав – и большим числом сезонных аспектов: с начала весны до осени их насчитывают до одиннадцати. Общая картина чередования весеннелетних аспектов в луговой степи изображена на рис. 9.122.
Луговые степи необыкновенно красочны. В течение веге-
тационного периода они постоянно изменяют свой облик. Ранней весной степь бурая, но уже в апреле, как по волшебству, раскрывает нежные лиловые бутоны сон-тра- ва, за ней спешит адонис весенний. Его крупные солнечно-желтые цветки появляются еще до развития листьев. Во время цветения адониса степь постепенно одевается в зеленый наряд. Зацветают незабудки, ирис безлистный, ветреница лесная. Начало лета знаменует цветение лугового шалфея. Наиболее красочной бывает степь в это время. Но вот зацветает перистый ковыль. Все остальные цветущие виды разнотравья скрываются под его пушистыми перьями. В это время степь особенно прекрасна, похожа на седое бескрайнее море. Проходят дни, постепенно степь одевается ярко-желтыми соцветиями подмаренника. Живокость клиновидная и чемерица черная – последние цветущие растения уходящего лета.
Животный мир евразиатских степей так же, как растительность, обладает своими подзональными и провинциальными особенностями. Например, различия в фауне Европейско-Казахстанских и Забайкальских степей можно увидеть, сравнив рис. 9.123 и 9.124.
Ограничимся краткой характеристикой животного мира Европейско-Казахстанской степной провинции. Ее границы хорошо очерчивают ареалы байбака и большого туш-
канчика (рис. 9.125). Фауна провинции представляет собой пестрый комплекс, переходный от биомов широколиственного леса к азиатским пустыням. Тем не менее, он имеет эндемичное ядро, в состав которого входят несколько видов сусликов, байбак, слепыш, большой тушканчик, обыкновенный хомяк, степная мышовка. К этой же группе относится антилопа сайгак. Из наиболее характерных птиц следует назвать стрепета, дрофу, малого журавля, степного луня, степного орла; из пресмыкающихся степную гадюку, разноцветную ящурку.
Для естественных ландшафтов открытых степных равнин характерны крупные стадные копытные и грызуны. Отсутствие естественных укрытий обусловило то, что степные копытные обладают быстрым бегом и хорошим зрением, а грызуны укрываются в норах. Суровая зима заставляет многих зверей и птиц откочевывать в более южные районы.
Следует подчеркнуть, что названные группы животных не только потребляют растительную массу, но и активно воздействуют на почвенно-растительный покров, разбивая дернину и роя многочисленные норы. Высокая биопродуктивность растительности составляет основу цепей питания в степных экосистемах (рис. 9.126).
Наблюдения в заповедниках показали, что умеренные нарушения являются необходимыми для устойчивого развития степ-
234
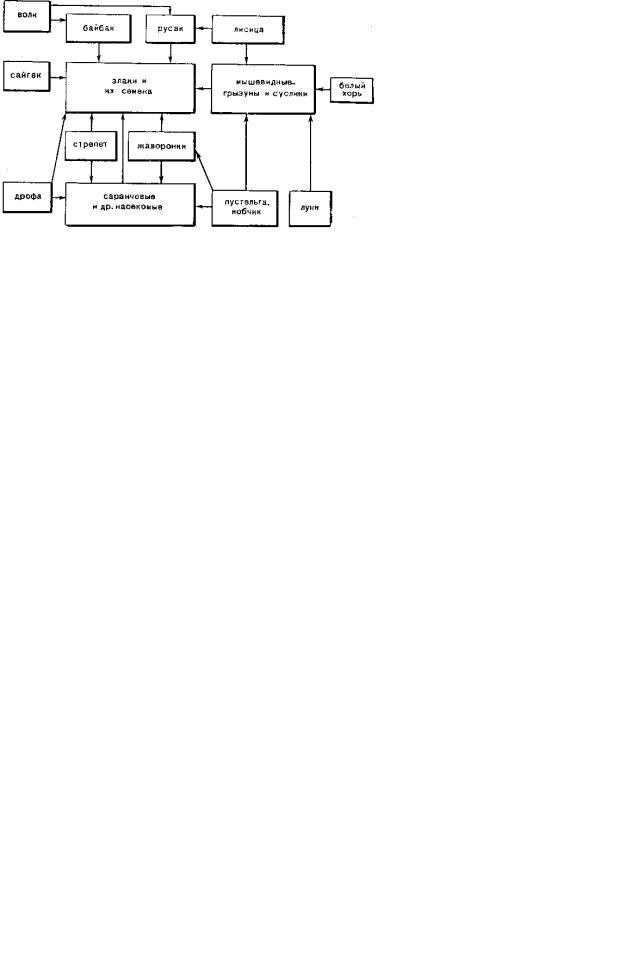
Рис. 9.126. Цепи питания в степных экосистемах, по Н. А. Бобринскому
ных сообществ: благодаря им появляются открытые участки по- чвы, доступные для семенного возобновления. Если же биопродуктивность растений не регулируется, накапливается мощный слой ветоши, дернины злаков уплотняются, так что семенное возобновление становится невозможным, многие виды не выдерживают конкуренции и биоразнообразие сообществ резко падает. Следует сказать, что в естественных условиях численность животных и биопродуктивность растительности саморегулируются и степные биомы развиваются устой-чиво.
Степные биомы Европейско-Ка- захстанских степей испытали особенно сильное воздействие человека. В Восточной Европе практически все степи распаханы, рассечены густой сетью дорог, заняты городами и промышленными предприятиями. Вместе с растительным покровом исчезли
Рис. 9.128. Ареалы прерий, по Т. Н. Гордеевой и О. С. Стрелковой:
Прерии: 1 – настоящая высокотравная восточная и южная; 2 – низкотравная и разнотравная; 3 – северные из овсяницы (Festuca); 4 – северо-западная; 5 – пустыни и полупустыни; 6 – территории с преобладанием засоленных почв, растительный покров несомкнутый
многие виды растений и животных, у других резко сократились ареалы.
Неарктическая область прерий Голарктического царства в
Северной Америке располагается вдали от атлантического океанического сектора, занятого лесными биомами (см. рис. 9.110). Данные об изменениях климата, растительности и почв прерий с востока на запад к подножью Скалистых гор отображены на профиле (рис. 9.127).
235

Доминантами растительного покрова являются дерновинные злаки, на востоке – высокие, а на сухом западе – низкие. Общая картина ареалов прерий изображена на карте (рис. 9.128).
В высокозлаковой или настоящей прерии господствуют дерновинные злаки, образующие травостой высотой 0,5 – 1 м. К числу доминантов относятся: ковыль (Stipa spartea),овсяница (Festuca scabrella), житняк (Agropyrum smithii), бородач, или грама
(Bouteloua curtipendula), тонконог (Koeleria
Рис. 9.130. Стадо степных бизонов
cristata); обильно разнотравье. Эти прерии почти сплошь распаханы.
Низкозлаковая прерия покрывает пространства Великих равнин. Здесь абсолютное господство переходит к плотнодерновинным злакам: бизоновой траве (Buchlo¸ dactyloides), траве грама (Bouteloua gracilis) (рис. 9.129). Эти прерии используются под пастбища или орошаемое земледелие.
Рис. 9.129. Доминанты низкозлаковых прерий:
а – бизонова трава (Buchlo¸ dactyloides); б – трава грама (Bouteloua gracilis)
Прерии относятся к обширной Сонорской области Неарктики, включающей широколиственные леса и субтропические биомы (см. рис. 9.68). Символом животного мира мог бы служить степной бизон, поголовье которого до колонизации Северной Америки европейцами, вероятно, достигало 50 – 60 млн. (рис. 9.130). Индейцы не могли нанести ему существенного урона, так как охотились с копьями или луком и стрелами. С 1872 по 1874 год белыми было убито более 5 млн бизонов, а к концу 1889 г. их осталось всего около тысячи. Для степного бизона роковой оказалась постройка транс-
Рис. 9.131. Охота на бизонов с поезда трансконтинентальной железной дороги в конце XIX века
континентальной железной дороги. Отстрел животных происходил прямо с проходящих поездов (рис. 9.131).
Южное полушарие.Основные площади злаковников располагаются в Южной Америки и Новой Зеландии.
Пампа – южноамериканский вариант субтропических степей занимает равнины Параны – Ла-Платы (рис. 9.132). Рельеф плоский, равнинный.
Климат субтропический. Зима теплая, температура самого холодного месяца 8 – 12 îС; летние температуры 22 – 25 îС. Годовое количество осадков возрастает от 500
– 600 мм на западе до 1000 мм у атланти- ческого побережья. Испаряется влаги несколько больше, чем выпадает. Два-три раза в течение каждых 10 лет случаются очень засушливые годы. Основная почвообразующая порода в пампе – л¸сс. На л¸се формируются черноземовидные почвы.
Безлесие пампы – явление во многом вторичное, возникшее еще до прихода европейцев, в результате выжигания растительности индейцами. К причинам безле-
236
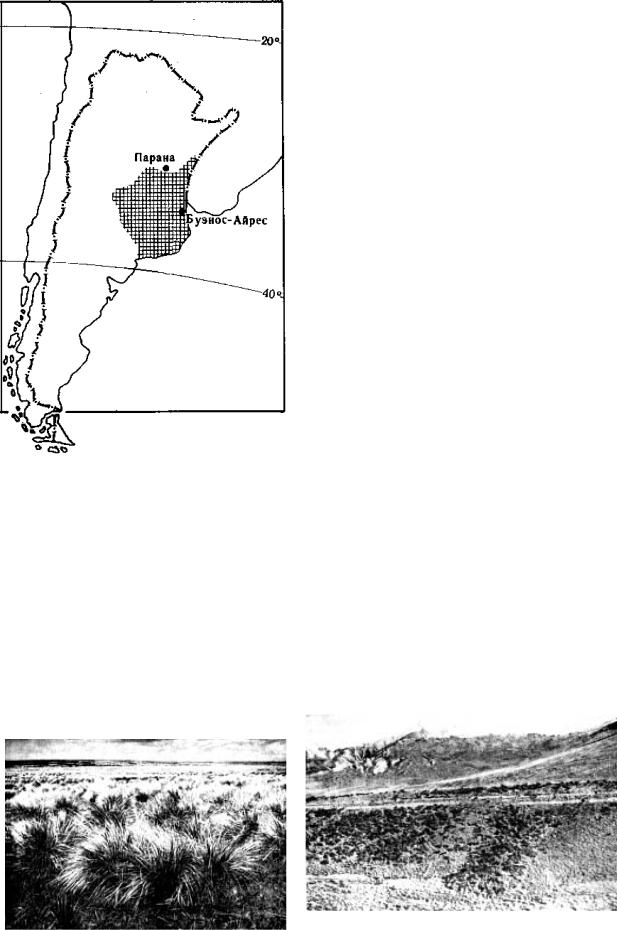
Рис. 9.132. Ареал пампы в Южной Америке, по Г. Вальтеру
сия относятся также периодические засухи, слабая дренированность и засоление грунтов.
В настоящее время - это наиболее густо населенные сельскохозяйственные районы Южной Америки. Первичная растительность пампы практически уже не существует. Однако в результате ее реконструкции установлено, что доминантами являлись злаки, многие рода которых характерны для лугов, степей и прерий Северного полушария: ковыль (Stipa), мятлик (Poa),
Рис. 9.133. Пампа с ковылем (Stipa brachychaeta), по Г. Вальтеру
аристида (Aristida), перловник (Melica), костер (Bromus) и др. (рис. 9.133); кроме того, для пампы характерны свои родаPaspalum,
Ischaemum, Cortaderia, Gynerium и др. Разнотравье представлено лупином (Lupinus), горошком (Vicia), вербеной (Verbena), портулаком (Portulaca), видами паслена.
Фауна пампы относится к Патагонско- Чилийской области Неотропического царства (см. рис. 9.113). В прошлом здесь обитали стада пампасского оленя и гуанако. В настоящее время многочисленны лишь грызуны - вискаша; встречаются броненосец, страус нанду.
В Новой Зеландии злаковники известны под названием туссок. Они распространены на Южном острове в дождевой тени Южных Альп от морского побережья до высоты 1500 м. Климат умеренно-теплый. Годовое количество осадков 600 – 500 мм. Почвы буро-серые.
Естественный растительный покров представлен злаковыми кочкарниками. Доминирующие рода – овсяница (Festuca, F. novae-zelandiae), мятлик (Poa, P. caespitosa), житняк (Agropyrum, A. scabrum) – характерны для степей Голарктического царства, т. е. имеют биполярный ареал. Это так называемый низкий туссок (рис. 9.134). Всего в группу формаций низких злаковников входит около 40 видов злаков и 35 видов сложноцветных. На влажных торфянистых почвах произрастает высокий туссок, образованый дантонией (Danthonia raoulii). Флористически эта формация беднее, так как высокие и плотно расположенные дернины дантонии подавляют другие виды.
Рис. 9.134. Туссоковый злаковник на о. Южный, Новая Зеландия, по Г. Вальтеру:
Доминирует овсяница (Festuca novae-zelandicae)
237
Со времен первых переселенцев злаковники Южного острова использовались как пастбища, поэтому девственных, нетронутых европейцами и домашними животными злаковых биомов не сохранилось.
9.2.6.Внетропические пустыни
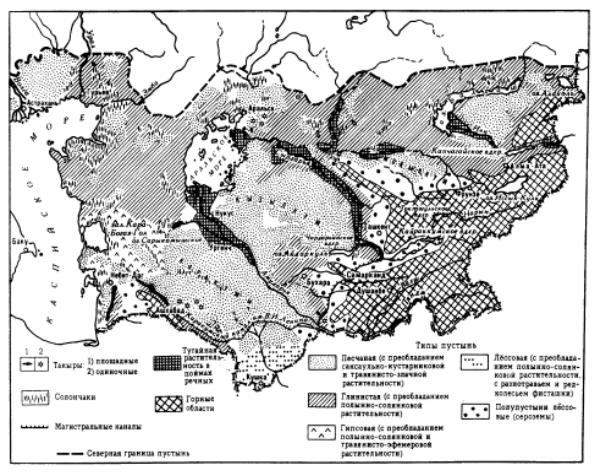
Âконтинентальном секторе Евразии к юго-востоку от зоны степей располагаются пустыни. Они занимают обширные пространства умеренного пояса Средней и Центральной Азии (рис. 9.135). На юге они смыкаются с пустынями жаркого пояса.
Переход от пустынь жаркого пояса к внетропическим пустыням происходит постепенно. Он связан прежде всего с понижением зимних температур. Устойчивый морозный период определяет своеобразие биомов. Границей, разделяющей тропические и внетропические пустыни, могут служить пределы возделывания финиковой пальмы, которая не переносит длительных заморозков.
Общие сведения о рельефе пустынь, формировании пустынных биомов, адаптациях растений и животных к жизни в условиях высоких температур и дефицита влаги рассмотрены при описании пустынь жаркого пояса (см. раздел 9.1.3). Остановимся на некоторых особенностях биомов пустынь Средней Азии и Казахстана.
Южные степи и северные пустыни объединены полосой динамического контакта, выделяемой в качестве полупустыни. По мере продвижения на юг дерновинные злаки постепенно уступают место пустынным полукустарникам: видам полыни (род
Artemisia, подроды Seriphidium и Dracunculus), прутняку (Kochia prostrata), разнообразным солянкам и т. п. Следует под- черкнуть, что наряду с полынью семейство маревые, или лебедовые (Chenopo-diaceae), куда входят все виды, объединяемые термином “солянки”, является ландшафтообразующим. Ареал саксаула (род Haloxylon, сем.
Chenopodiaceae) довольно точно очерчивает границы пустынь Средней Азии и Казахстана (рис. 9.136).
Пустынные ландшафты Туранской низменности простираются на 10 – 12î по широте. На этом протяжении зональные изменения термических условий и режима увлажнения приводят к формированию двух подзон – северных и южных пустынь. Климатическими условиями
образования внетропических пустынных биомов, общими для пустынь жаркого пояса являются: большая солнечная радиация, исключительная засушливость всего вегетационного периода, высокие летние температуры (25 – 30 îС), малое количество ат-
Рис. 9.136. Ареал рода саксаул, по “Жизнь растений”
238
мосферных осадков (в среднем 100 – |
ной стадии. Значительное участие в почво- |
150 ìì). |
образовании принимают водорастворимые |
Испаряемость в 10 – 25 раз превышает |
соли – карбонаты, сульфаты и поваренная |
сумму выпадающих осадков. Зимние темпе- |
соль. Они переходят в почву из сильно ми- |
ратуры отрицательные -10, -15 îÑ, ëèøü íà |
нерализованных материнских пород или |
юге Туранской низменности они несколько |
поднимаются по капиллярам из грунтовых |
âûøå 0 îС; абсолютные минимумы на се- |
вод. В то же время в пустынных почвах |
вере Казахстана достигают -50 îÑ, â Òàø- |
очень мало гумуса (менее 1 %). Это связано |
кенте -30 îÑ. |
с тем, что растительность бедна, и в почву |
В жизни пустынь в условиях дефицита |
поступает небольшое количество органи- |
атмосферных осадков большое значение |
ческих остатков, которые к тому же быстро |
имеют подземные воды, в частности грун- |
минерализуются. Пестрота почво-грунтов, |
товая верховодка. В пустынях, сформиро- |
разная степень засоления, условия увлаж- |
ванных на месте древних аллювиальных |
нения– причины комплексности раститель- |
равнин, они образуют сплошное зеркало. |
ного покрова. |
Грунтовые воды засолены, часто залегают |
В основе классификации пустынь |
неглубоко (3 - 4 м) и доступны для корней |
Средней Азии и Казахстана лежит характер |
растений; местами располагаются линзы |
поверхностных отложений и почвогрунтов. |
пресной верховодки. |
Основными типами пустынь являются: пес- |
Засушливый климат обусловил особый |
чаные, глинистые, гипсовые, лессовые; сво- |
тип почвообразования: для южных пустынь |
еобразные природные комплексы представ- |
характерны различного типа сероземы, для |
лены такырами, солончаками и поймами |
северных – серо-бурые почвы. На огромных |
рек; особый тип ландшафтов связан с гора- |
территориях песчаных пустынь почвообра- |
ìè (ðèñ. 9.137). |
зовательный процесс находится в началь- |
|
Рис. 9.137. Пустыни Средней Азии и Казахстана, по: “Пустыни”
239

В тесной связи с почвогрунтами нахо- |
На нем отчетливо видны гряды песков, |
|
дится растительность. Выделяются следую- |
заросшие псаммофитами, и солончаковая |
|
щие типы пустынной растительности (вне |
депрессия с сообществами галофитов. |
|
речных пойм): псаммофитный древесно- |
Фауна пустынь Средней Азии и Казах- |
|
кустарниковый на песках (Haloxylon |
стана относится к Арало-Каспийской обла- |
|
persicum, H. aphyllum, Calligonum caput- |
сти Палеарктики (рис. 9.139). Она отлича- |
|
medusae, C. setosum, Salsola richteri, |
ется своими специфическими особенностя- |
|
Ammodendron conollyi, Ephedra strobilacea è |
ми и вместе с тем имеет переходный харак- |
|
др.); гипсофильный мелкокустарничковый |
тер между сахара-средиземноморской и цен- |
|
на щебнистых и каменистых третичных |
тральноазиатской фаунами. |
|
плато (Salsola arbuscula, S. rigida, S. |
Адаптации растений и животных к |
|
gemmascens, S. laricifolia, Anabasis salsa, |
жизни в условиях пустыни вырабатывались |
|
Hammada leptoclada); эфемерово-эфемеро- |
на протяжении более 100 млн. лет, с мело- |
|
идный на л¸ссах; полукустарниковый по- |
вого и третичного периодов. Они шли по |
|
лынный на суглинках (Artemisia semiarida, |
пути приспособления к засухе, жаре, боль- |
|
A. kemrudica, Kochia prostrata, Eurotia |
шой амплитуде температур, бедным и засо- |
|
ceratoides с участием многолетних злаков, а |
ленным грунтам, к сыпучим пескам и |
ò. |
также эфемеров и эфемероидов); водорос- |
п. Главное направление адаптаций у расте- |
|
лево-лишайниковый на такырах со своеоб- |
ний привело к образованию многочислен- |
|
разными группировками пустынных ли- |
ной группы ксерофитов (см. табл. 9.2). |
|
шайников и синезеленых водорослей (Пет- |
Основными приспособительными |
|
ðîâ, 1973) . |
признаками ксерофитов являются особен- |
|
В качестве иллюстрации зависимости |
ности формирования их листа, ведущие к |
|
растительности от почвогрунтов приведен |
уменьшению размеров листовой пластин- |
|
аэрофотоснимок песчаной пустыни Караку- |
ки – микрофилии и афилии (Василевская, |
|
ìû (ðèñ. 9.138). |
1954). Продукция надземной части пустын- |
|
|
ных растений ничтожна по сравнению с |
|
|
продукцией подземной: на корневую систе- |
|
|
му приходится до 95% всей фитомассы. Кор- |
|
|
ни очень глубоко проникают в землю, дос- |
|
|
тигая капиллярной каймы грунтовых вод на |
|
глубине до 20 – 25 м.
Весьма своеобразны приспособления псаммофитов – растений, живущих на рыхлых, иногда даже подвижных песках. Их корни протягиваются по склонам барханных цепей на 25 – 30 м и уходят в глубину на 2- 2,5 м, достигая капиллярного подпора грунтовых вод и обеспечивая растения достаточным количеством влаги. Во время передвижения песков ветер выдувает из-под корней грунт, они частично обнажаются, но не гибнут, так как хорошо защищены от палящих лучей солнца. Засыпанные песком кусты не погибают. Они дают массу придаточ- ных корней, способствующих закреплению песка (рис. 9.140).
Плоды и семена многих псаммофитов
Рис. 9.138. Грядовый рельеф песков с сообществами псам- имеют приспособления для переноса вет-
мофитов; серое овальное пятно (а) – солончаковая деп- ром (рис. 9.141). У песчаной акации они рессия с сообществами галофитов. Аэрофотоснимок
240