

80. Обмен железа: всасывание, транспорт, депонирование, регуляция, Роль железа в организме. Нарушения обмена железа в организме человека.

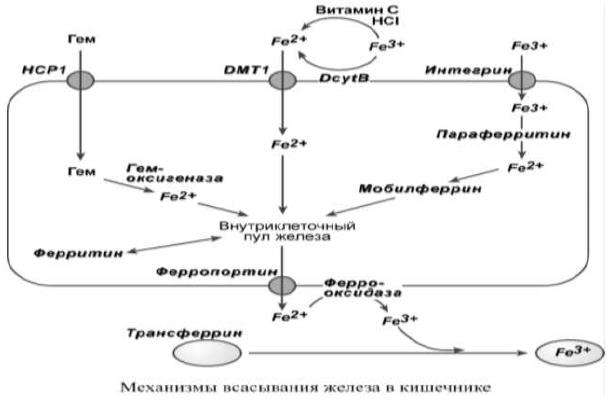
Всасывание происходит в проксимальном отделе тонкого кишечника в количестве около 1,0-2,0 мг/день (10-15% пищевого железа). При этом железо должно быть в виде двухвалентного иона, в то же время с пищей поступает преимущественно трехвалентное железо. Только железо мясных
продуктов находится в гемовой форме (Fe2+), и поэтому хорошо всасывается.
Существуют три способа перемещения железа из просвета кишечника в энтероциты:
Негемовое железо (III) захватывается интегрином, восстанавливается до Fe (II) параферритином, и при помощи мобилферрина перемещается в клетке, активность и роль этого пути низкая.
Негемовое железо (III) восстанавливается до Fe (II) при помощи аскорбиновой кислоты, соляной кислоты или при участии ферроредуктазы (DcytB, дуоденальный цитохром B) и далее переносится внутрь белком DMT-1 (divalent metal ion transporter-1).
Гемовое железо связывается с белком НСР1 (heme carrier protein 1), в цитозоле высвобождается из гема при действии гемоксигеназы и далее переносится по клетке.
Транспорт железа

Холотрансферрин – белок, который содержит ионы Fe3+
Апотрансферрин – белок, которой не содержит ионы Fe3+ (свободный)
После всасывания железо либо откладывается в клетках кишечника в составе ферритина, либо попадает в кровоток и в комплексе с трансферрином переносится в клетки печени, костного мозга или других тканей.
Холотрансферрин связывается со своим трансферриновым рецептором, образуя тройной комплекс (железо, апотрансферрин и рецептор). Этот комплекс в составе эндосомы перемещается в цитоплазму. Внутри эндосомы в результате работы H+-АТФ-азы создаётся кислый рН, из-за чего железо диссоциирует из комплекса и поступает в цитоплазму, составляя свободный пул. Далее освободившееся железо может мобилферрином доставляться к месту использования, например, встраиваться в порфириновое кольцо гема, включаться в железопротеины или депонироваться с ферритином.
Эндосома, содержащая апотрансферрин-рецепторный комплекс, возвращается обратно к плазматической мембране. При нейтральных рН апотрансферрин имеет низкое сродство к своему рецептору и высвобождается в кровоток, что позволяет другой молекуле холотрансферрина связаться с рецептором.
Вне связи с белками железо очень токсично, так как запускает свободно- радикальные реакции с образованием активных форм кислорода.


Регуляция
Эритропоэтин стимулирует эритропоэз и активирует механизмы, повышающие абсорбцию железа
Гепсидин ингибирует абсорбцию и реутилизацию (т.е. повторное использование) железа
Механизм действия гепсидина: снижает активность белка ферропортина, отвечающего за выход ионов железа из клетки в кровь (в кишечнике), а также ионы железа задерживаются в макрофагах.

Основой регуляции является система IRE/IRP (железочувствительный элемент/железочувствительный белок), которая обеспечивает регуляцию:
o синтеза рецепторов к трансферрину,
o синтеза ферритина.
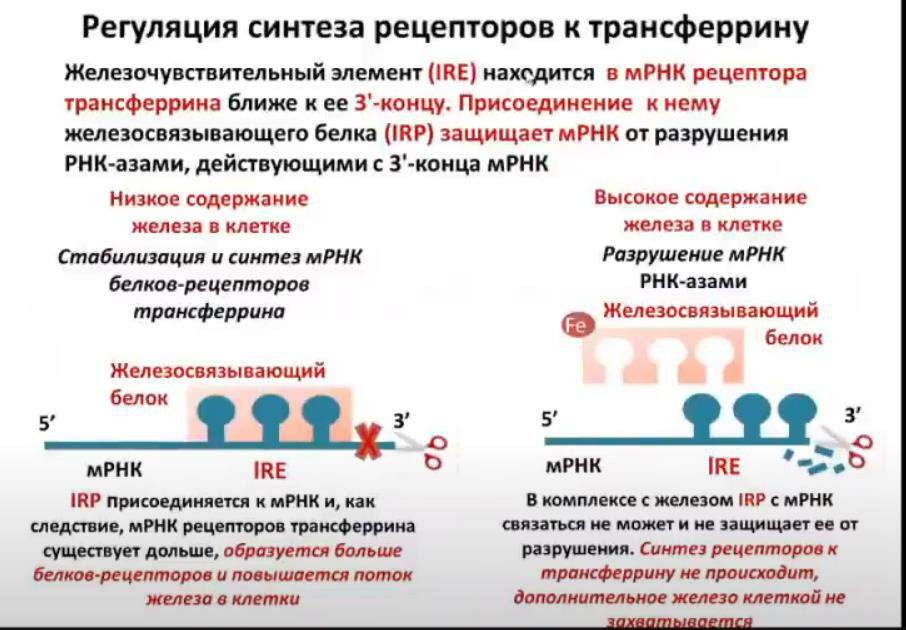

Присоединение активного IRP к 5'-концу мРНК ферритина (при низком уровне железа) не позволяет ей участвовать в процессе трансляции и синтезе новых молекул ферритина. Когда концентрация железа в клетке возрастает, оно присоединяется к IRP и снижает его сродство к матричной РНК. Конечным результатом является увеличение синтеза молекул ферритина, депонирующего железо.