Роль железа в организме:
Обратимое связывание, транспорт и сохранение кислорода (гемоглобин и миоглобин)
Перенос электронов (железосерные белки, цитохромы а,b, с, цитохром с оксидаза)
Функционирование активных центров окислительно- восстановительных ферментов
окисление биомолекул (цитР450, пероксидазы)
разрушение (каталаса, супероксиддисмутаза) и генерирование активных форм кислорода (НАДФН-оксидаза)
Избыток железа
Существует аутосомно-рецессивное заболевание гемохроматоз, связанное с нарушением гепсидиновой регуляции и избыточным всасыванием железа в кишечнике. В результате железо накапливается в органах и тканях.
Общее содержание железа в организме достигает 20-60 г при
норме 2-4 г. Постепенно развиваются цирроз печени, кардиомиопатии, сахарный диабет 1 типа, артрит.
Железодефицит
Причинами железодефицита являются недостаток его в пище,
заболевания ЖКТ со снижением всасывания (гастриты), потери железа с кровью при менструальных, кишечных или иных кровотечениях
При недостаточности железа в организме мобилизация резервов происходит в следующем порядке:
железо из депо (ферритин),
затем в клетках (кроме эритроидных) снижается количество гемопротеинов до жизнеспособного минимума,
далее истощаются запасы сывороточного железа (холотрансферрин),
в последнюю очередь страдает синтез гемоглобина.
Таким образом, железодефицитная анемия является проявлением
крайнего дефицита железа, и нормальная концентрация гемоглобина крови не должна быть критерием обеспеченности организма железом.
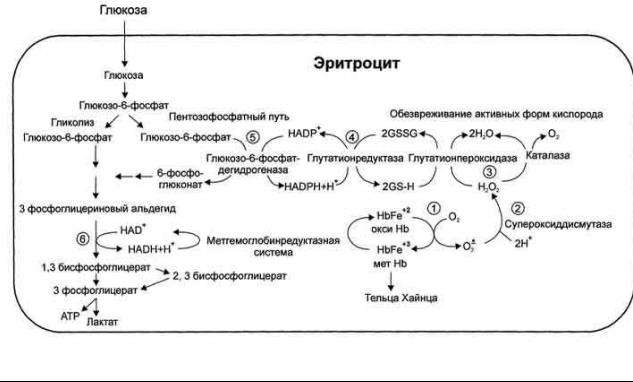
Особенности метаболизма в эритроцитах. Механизмы свободно- радикального окисления (СРО) и антиоксидантная защита в эритроцитах (АОЗ).
Зрелые эритроциты лишены ядра, поэтому в клетке не синтезируются белки.
Эритроциты не имеют митохондрий, поэтому в клетке не протекают реакции ЦТК, ЦПЭ, β- окисления жирных кислот.
Основным энергетическим субстратом эритроцита является глюкоза – лактат в кровь.
Энергия, поставляемая гликолизом, обеспечивает поддержание целостности плазматической мембраны и работу Na+, K+-АТФазы.
Присутствие в них фермента дифосфоглицератмутазы. Дифосфоглицератмутаза катализирует образование 2,3- дифосфоглицерата из 1,3-дифосфоглицерата.
.Образующийся только в эритроцитах 2,3-дифосфоглицерат служит аллостерическим регулятором связывания О2 гемоглобином.
В реакции дегидрирования глицеральдегид-3-фосфата в гликолизе образуется НАДН. Этот кофермент является кофактором фермента метгемоглобинредуктазы, участвующей в восстановлении метгемоглобина в гемоглобин по следующей схеме:
![]()
10 % глюкозы распадается в эритроците в пентозофосфатном пути. Образующийся при этом НАДФН обеспечивает восстановление глутатиона и поддерживает его оптимальную концентрацию. Восстановленный глутатион необходим для поддержания в
восстановленной форме SH-групп белков; препятствует окислению гемоглобина; предотвращает перекисное окисление липидов мембран.
Защита от СРО:
Эритроциты содержат ферментативную систему, предотвращающую токсическое действие активных форм кислорода и разрушение мембран эритроцитов.
Супероксидный анион супероксиддисмутазой превращается в пероксид водорода: O2- + O2- + Н+
→ H2О2 + O2.
Пероксид водорода под действием каталазы (2Н2О → 2Н2О + О2) или глутатионпероксидазы
(2GSH + 2Н2О2 → GSSG + 2Н2О) превращается в Н2О и О2. Донором водорода для глутатионпероксидазы является восстановленный глутатион (GSH).
Окисленный глутатион (GSSG) восстанавливается ферментом глутатионредуктазой, кофермент которого NADPН образуется в пентозофосфатном пути катаболизма глюкозы.
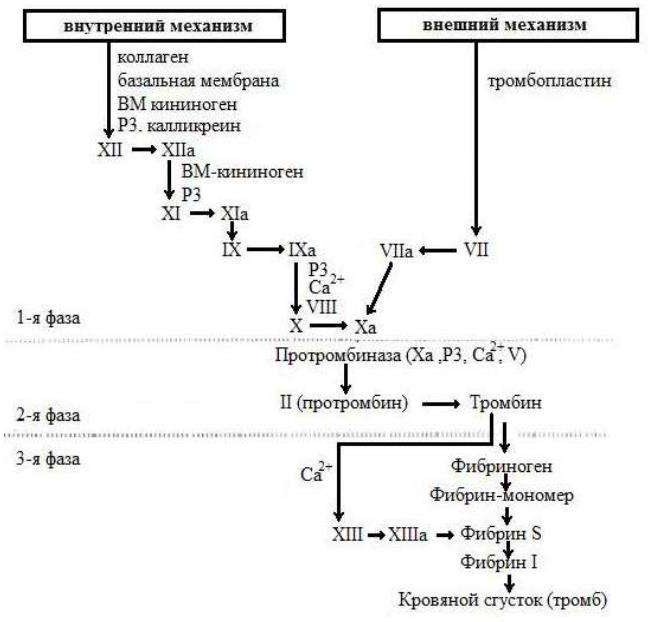
 Внешний
и внутренний пути свертывания крови.
Каскад реакций
прокоагулянтного
этапа свертывания крови. Механизмы
активации
ферментов
свертывающей
системы
крови.
Витамин
К
и
его
антагонисты
(дикумарол).
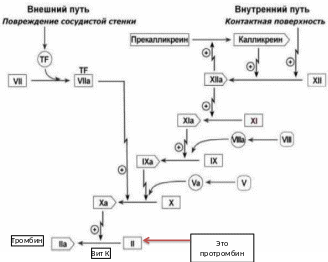
Внешний
и внутренний пути свертывания крови.
Каскад реакций
прокоагулянтного
этапа свертывания крови. Механизмы
активации
ферментов
свертывающей
системы
крови.
Витамин
К
и
его
антагонисты
(дикумарол).
TF – тканевый фактор.
Ферменты активируются каскадно за счет постепенного присоединения к неактивной форме кофактора (TF,калликреин, Va, Xа и тд).
В печени при участии витамина К происходит синтез его неактивного предшественника –
протромбина, который в дальнейшем циркулирует в плазме. В плазме крови превращение
протромбина в тромбин происходит непосредственно под действием фактора Xa (совместно с Va). Дикумарол блокирует переход витамина К в активную форму.
На всякий случай