

Толстые нити образованы белком миозином
Тонкие нити образованы актином,тропомиозином и тропонином
Биохимические механизмы мышечного сокращения и расслабления. Роль градиента одновалентных ионов и ионов кальция в регуляции мышечного сокращения. Особенности сокращения гладких мышц.
Этапы мышечного сокращения
l –й этап. Мышечное сокращение начинается с нервного импульса. Под воздействием ацетилхолина развивается возбуждение клеточной мембраны и резко повышается ее проницаемость для ионов Са2+.
Это приводит к выходу ионов Са2+ из саркоплазматического
ретикулума в цитозоль. Содержание ионов Са2+ в цитозоле миофибриллы увеличивается до 10-5 моль/л.
–й этап. Ионы Са2+ связываются с С-субъединицей тропонина, что вызывает конформационные изменения тропонина и тропомиозина.
В результате на актине открываются центры связывания головок миозина
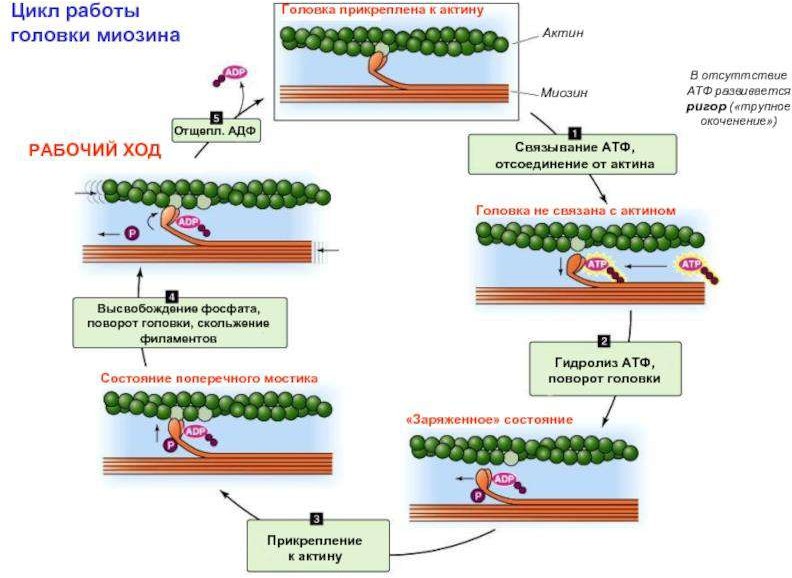
– Й этап.
Фиксация АТФ на головке миозина
Гидролиз АТФ
Продукты гидролиза (АДФ и Ф) остаются фиксированными, а выделившаяся энергия аккумулируется в головке.
Содержащая АДФ и Фн миозиновая «головка» присоединяется к центру связывания на актине под углом 90°, образуется актин- миозиновый комплекс
- й этап. АДФ и Рн выходят из актин-миозинового комплекса. Происходит спонтанный поворот миозиновой «головки» на 45°
При этом тонкая актиновая нить движется вдоль миозиновой в направлении к М-линии. Происходит сокращение саркомера.
Головки миозина "работают" циклично, как весла у лодки, поэтому этот процесс называется "весельным механизмом" мышечного сокращения. Комплекс "актин-миозин«очень прочен. Он может быть разрушен только при сорбции новой молекулы АТФ.
й этап. Новая молекула АТФ связывается с «головкой» миозина, что приводит к распаду актомиозина.
Связывание (но не гидролиз) АТФ с миозином сопровождается отделением миозина от актина и создает возможность повторения цикла поперечных мостиков.
На этой стадии атф не расщепляется, т.Е. Служит не источником энергии, а аллостерически изменяет конформацию миозиновой головки и тем самым ослабляет связь миозина с актином
При поступлении нового нервного импульса к мышце цикл сокращения повторяется
Мышечное расслабление
в отсутствии нервного импульса содержание Са2+ в саркоплазме падает ниже 10-7 моль/л вследствие его поглощения саркоплазматическим ретикулумом;
распад комплекса ТnС•Са2+
изменение конформации тропонина I и тропомиозина приводит к закрытию активных центров F-актина и ингибирование дальнейшее
взаимодействие миозиновой головки с F-актином
миозиновые головки в присутствии АТФ отделяются от F-актина, вызывая расслабление.
Не только процесс сокращения, но и процесс расслабления нуждается
в АТФ, потому что если нет АТФ, то не работает Са2+-зависимая АТФаза. В этих условиях кальций связан с тропонином "С" - вся система находится в активном состоянии, нет распада актомиозинового комплекса - мышца постоянно находится в состоянии сокращения. Такая ситуация наблюдается после смерти организма в состоянии "трупного окоченения".
В сердечной мышце основным источником ионов Са2+ для возбуждения служит внеклеточная жидкость, при отсутствии Са2+ во внеклеточной жидкости сокращения сердечной мышцы прекращаются в течение одной минуты. Скелетная мышца в таких условиях может сокращаться часами.