

1.2 Эмбриональное развитие
Эмбриональный период характеризуется эндогенным питанием зародыша за счет запасов желтка в яйце, полученных от материнского организма, составляющих у рыб от 25 до 40% от веса тела самки. Развитие зародыша в период эмбриогенеза происходит внутри яйцевой оболочки и в таком состоянии он называется эмбрионом. С момента вылупления из оболочки при развитии в свободном состоянии он принимает название предличинки или свободного эмбриона.
I этап — оводнение икринки, образование перивителлиновой полости, образование бластодиска.
Возраст 1–3 мин — начало оводнения (набухания) икринки. Набухание икры и отставание первичной оболочки — зоны радиаты от поверхности желтка начинается сразу после попадания икры в воду. При этом толщина оболочек за счёт набухания увеличивается. Механизм образования перивителлинового пространства обусловлен выделением под оболочку содержимого кортикальных гранул. Это способствует активному поступлению воды под оболочку. Вторичная оболочка, образованная остатками отростков клеток фоликулярной оболочки — хориона, также набухает, образуя выросты. На рис. 15 показано, что одновременно с образованием перивителлинового пространства и концентрацией плазмы на анимальном полюсе, яйцо поворачивается этим полюсом вверх в наибольшую часть «околожелточной полости» за счет гидростатических свойств жировых капель, в определённой степени сопряжённых с анимальным полюсом. [15]
К моменту завершения кортикальной реакции часть цитоплазмы уже собирается на анимальном полюсе. Параллельно с этим процессом происходит формирование зиготы: самого зародыша. Головка спермия, проникшая в поверхностный слой ооплазмы анимального полюса ооцита, провоцирует второе редукционное деление. Редукционное тельце удаляется из ооплазмы в перивителлиновое пространство, а головка спермия продолжает постепенно погружаться в яйцеклетку. По В. В. Махотину (1982) второе деление созревания и выброс редукционного тельца у зародышей трески происходит за один час. Заканчивается данный этап оплодотворения расположением женского и мужского пронуклеусов друг против друга, т. е. преобразованием яйцеклетки в функциональную зиготу. [31]
Возраст 4 ч — окончание набухания икры (рис. 15, а). Перивителлиновая полость между яйцевой оболочкой и поверхностью яйца достигла максимального размера. Расчеты показывают, что перивителлиновая полость составляет к концу набухания треть объема икринки. После оплодотворения свободная от желтка цитоплазма стягивается к анимальному полюсу, и первоначальное небольшое утолщение плазмы значительно увеличивается.
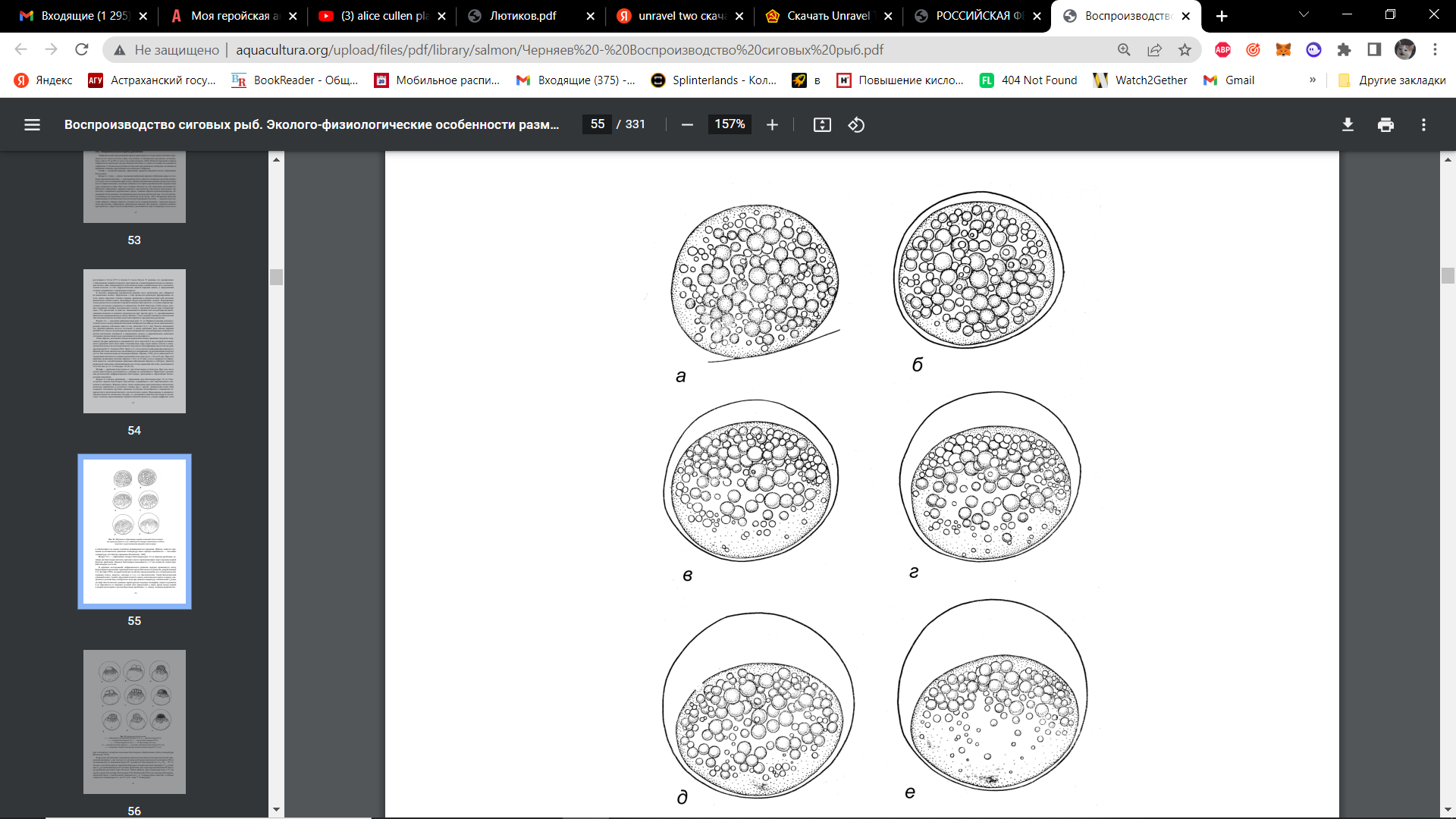
Рисунок 15. Набухание и образование перивителлиновой полости икры (на серии рисунков от а до е наблюдается поворот анимального полюса, заметного по расположению жировых капель вверх:
II этап — дробление бластодиска от двух бластомеров до бластулы. При этом число клеток (бластомеров) увеличивается, а размеры их уменьшаются. Происходит дальнейшая региональная дифференцировка бластомеров, приводящая к образованию билатеральной симметрии.
Возраст 8 ч. — образование двух бластомеров (рис. 16, б). Анимальный полюс яйца совершает медленные круговые движения вследствие волнообразного сокращения поверхностного цитоплазматического слоя желточного мешка. Перемещение в перивителлиновой жидкости плазменного бугорка, а в дальнейшем шапочки бластомеров способствует лучшему перемешиванию перивителлиновой жидкости, ускоряя диффузию газов и обеспечивая тем самым газообмен развивающегося зародыша. Причем, скорость вращения имеет прямую зависимость — чем выше температура, тем быстрее вращение. [40]
Возраст 12 ч — образование четырех бластомеров (рис. 16, в). Борозда дробления, делящая два бластомера пополам, проходит строго перпендикулярно через середину первой борозды дробления. Диаметр бластомеров уменьшает. [20]
В пределах оптимальных температур можно использовать более простой способ определения величины τ0 как частного от деления всей продолжительности интервала от осеменения до появления борозд IV деления (16 бластомеров) на 5, так как у костистых рыб до появления борозды I деления проходит примерно 2 τ0 и плюс еще 3 τ0 до появления борозд IV деления дробления.
Возраст 1 сут — образование восьми бластомеров (рис. 16, г). Диаметр бластомеров равен 0,4 мм, они расположены в один слой на анимальном полюсе.
Возраст 2 сут — образование 16 бластомеров (рис. 16, д), Борозды дробления направлены как горизонтально, так и перпендикулярно к вертикальной оси икринки. Вследствие этого часть бластомеров не соприкасается с желточным мешком.
Возраст 2,5 сут — образование 32 бластомеров (рис. 16, е). После пятого митотического цикла, во время которого перед появлением борозд дробления, адгезия мембран бластомеров слабеет, диаметр бластомеров уменьшился до 0,2 мм. Под анимальным полюсом происходит дальнейшее слияние жировых капель и их концентрация. Диаметр бластомеров уменьшается до 0,2 мм.
Возраст 4 сут — средняя морула (рис. 16, ж). Подсчет числа бластомеров становится затруднительным и для определения стадии развития приходится ориентироваться по размерам бластомеров. Диаметр бластомеров равен 0,1 мм.
Возраст 5,5 сут. — поздняя, или мелкоклеточная, морула (рис. 16, з). Бластомеры мелкие (диаметр около 0,03 мм). На седьмые сутки развития внутри морулы появляется темная зона уплотненных мелких бластомеров (рис. 16, и). На девятом митотическом цикле этапа дробления формируется нижний слой бластомеров зародышевого диска. Этот клеточный материал, освобождаясь от клеточных мембран, образует многоядерный синцитий, образуя базальный перибласт. Параллельно происходит дифференцировка покровного слоя клеток — перидермы. С появлением этих двух провизорных структур завершается этап дробления, занимаюшее порядка 8 суток, при температуре воды нерестилищ от +4º до +1ºС. [31]

Рисунок 16. Дробление бластодиска: а — образование плазменного бугорка (5 ч); б — два бластомера (8 ч); в — четыре бластомера (12 ч); г — восемь бластомеров (24 ч); д — 16 бластомеров (2 сут); е — 32 бластомера (2,5 сут); ж — крупноклеточная морула; з — поздняя, мелкоклеточная морула (5,5 сут); и — появление темной зоны внутри мелкоклеточной морулы (7,5 сут)
III этап — бластуляция. Возраст 8 сут — в области скопления мелких бластомеров, четко выделяющихся темным пятном внутри мелкоклеточной морулы, образуется полость бластоцеля. Овальная полость бластоцеля при боковом микроскопировании обнаруживается между перибластом и бластодермой в виде небольшой светлой линзы, расположенной эксцентрично по отношению к центру бластодиска (рис. 17, а). Полость бластоцеля костистых рыб образуется путем разрыхления, утраты межклеточной адгезии и размножения внутренних клеток в результате митотических делений [1].
IV этап — гаструляция. Глубокие клетки рассортировываются на эпибласт и гипобласт и занимают определенное положение: верхнюю часть зародышевого диска представляет эпибласт, а нижний пласт, представлен гипобластом. Таким образом, осуществляется процесс гаструляции. При дальнейшем развитии зародыша часть полости бластоцеля обособляется от зародышевого щитка и превращается в перибластический синус. Благодаря координированным центробежным миграциям клеток гипобласта формируется утолщенный край зародышевого диска — «зародышевое кольцо» и «краевой узелок». На этом процесс гаструляции у рыб с меробластическим типом дробления завершается. [30; 42]
Возраст 11 сут — начало эпиболии (рис. 17, б). Бластодерма начинает перемещаться по поверхности желточного мешка в сторону вегетативного полюса. Диаметр бластодиска увеличивается с 1,5 (на этапе бластулы) до 1,7 мм. Бластодерма неоднородна. В наиболее утолщенной краевой части бластодиска образуется краевой узелок (рис. 17, в). В этой области происходит активное деление клеток. Краевой узелок увеличивается в длину, расслаивается на экто-, энто- и мезодерму и принимает вид краевого язычка (рис. 17, д). Бластоцель заметно увеличивается, верхняя стенка его утончается и выпячивается. Такая модификация бластоцеля называется перибластическим синусом (рис. 17, е).

Рисунок 17. Образование зародыша-бластулы и начало обрастания желточного мешка: а — бластула (8 сут); б — увеличение полости бластоцеля и начало эпиболии (9 сут); в —образование краевого узелка (11 сут); г — 1/3 бластодермы покрывает желток (12 сут); д — образование краевого язычка (13 сут); е — половина желтка покрыта бластодермой зародыша (14 сут)
V этап — органогенез: образование нервной «трубки», хорды, мускулатуры и т. д.
Нарастающая краевая часть бластодиска окружена пояском краевой мезодермы, которая темноватой полоской охватывает прозрачную сферу желточного мешка.
Клетки гипобласта последовательно мигрируют из зародышевого кольца, достраивая каудальный конец зародыша, в результате чего тело удлиняется, превращаясь в зародышевую полоску (рис. 17, е).
Возраст 15 сут — начало сегментации мезодермы (рис. 18, а). Длина зародыша 2 мм, три четверти желточного мешка покрыты бластодермой. Головной отдел эмбриона расширен, видна нервная бороздка. Ближе к заднему концу тела зародыша образовался первый мезодермальный сегмент. Дальнейший рост эмбриона и появление новых сегментов сопровождаются обрастанием желточного мешка (рис. 18, б).
Сегментации, то есть расчленению на отдельные блоки — сомиты, подвергается лишь утолщенная дорсальная часть мезодермы. При этом происходит объединение мезодермальных клеток в группы, разделенные узкими промежутками. Число сомитов (сегментов) постепенно возрастает благодаря их обособлению в задней несегментированной части зародыша. На протяжении этапа происходит дифференциация сомитов. Сначала от него отделяется спланхнотом, и в области отделения от «ножек» сомита обособляется промежуточная мезодерма. Часть ее преобразуется в выделительную систему зародыша, называемую нефротомом. Самый передний отдел тяжа формирует предпочку — пронефрос, а остальной длинный отрезок до зачатка мочевого пузыря — предпочечный выводной канал. [30]
Другая часть промежуточной мезодермы в виде мезенхимных клеток мигрирует под хорду и сливается в продольный тяж клеток, образующий магистральные кровеносные сосуды: спинную аорту и задние кардинальные вены. Третья часть мезенхимных клеток представляет собой склеротомную мезенхиму. Ее клетки мигрируют к хорде и охрящевают впоследствии на ее оболочке в четырех местах — в месте каждого будущего позвонка. После отделения от сомитов нефро-, склеро-, дерматома, а также мезенхимы кровеносных сосудов остается их основная часть — миотомы, которые и образуют туловищную мускулатуру. [19]
Возраст 25–27 сут — желточная пробка. Нарастающая бластодерма постепенно покрывает все уменьшающуюся свободную часть желточного мешка, называемую желточной пробкой. Утолщенный головной отдел дифференцируется на передний и задний. В плотном зачатке переднего мозга формируется полость, и он становится выше остальных отделов. В зачатках глаз появляются полости, преобразующиеся в дальнейшем в глазные пузыри. На уровне первичного заднего мозга с вентральной стороны эмбриона в слое кишечной энтодермы формируются жаберные карманы». [31]
Возраст 35 сут — образование глазных пузырей (рис. 18, г). Желточная пробка полностью замыкается. В теле эмбриона 16 миотомов: под передним отделом головы, в котором сформированы энцефаломеры, закладывается сердечная трубка. По бокам головы образовались глазные пузыри, позади них на некотором удалении заложились слуховые плакоды — зачатки органов слуха и равновесия. На месте замыкания желточной пробки сохраняется затемненная зона с вкрапленными в нее мелкими жировыми каплями.

Рисунок 18. Органогенез, сопровождающийся обрастанием желточного мешка бластодермой и замыканием желточной пробки: а — начало сегментации мезодермы (появление первого сегмента, 15 сут); б — образование трех мезодермальных сегментов (20 сут); в‑девять сегментов (замыкание желточной пробки, 25–27 сут); г — образование глазных пузырей (35 сут); д — образование хрусталика (40 сут); е — образование сердечной трубки (45 сут)
VI этап — обособление хвостового отдела от желточного мешка. Возраст 40 суток — образование хрусталика в глазах эмбриона (рис. 18, д). Начинает интенсивно расти хвостовая почка эмбриона. В головном отделе обособились передний, средний и продолговатый отделы мозга. В глазах появился хрусталик. В переднем отделе головы, перед глазами, заложены обонятельные плакоды. Задняя часть хвоста начинает обособляться от желточного мешка.
VII этап — появление системы кровообращения. Возраст 45 сут — начало пульсации сердечной трубки (рис. 18, е). В результате небольших изгибов тела и хвоста центр тяжести зародыша смещается, что заставляет эмбрион медленно переваливаться с боку на бок внутри оболочки. В туловище эмбриона 40 миотомов. Под хордой закладывается кишечная трубка в виде продольного тяжа. Слуховая плакода, в которой появилась полость, превращается в слуховую капсулу. В передней части головного мозга появляется эпифиз, в задней сформировывается мозжечок. В области 7–8 миотомов по обеим сторонам тела образуются зачатки грудных плавников.
Возраст 55 сут — начало пигментации глаз (рис. 19, а). В глазах появился черный пигмент — меланин. В теле эмбриона 60 сегментов. Пульсирующая сердечная трубка изогнута под прямым углом, передняя часть ее представляет собой зачаток желудочка, задняя — предсердия. Под кишечником в области грудных плавников начинает обособляться печень. В задней части кишечника образуется анальное отверстие. Зачатки грудных плавников покрыты крупными одноклеточными железами. Тело и хвостовая часть эмбриона слабо изгибаются.
Возраст 65–70 сут — начало образования форменных элементов крови (рис. 19, б). На этом этапе развития сердце пульсирует еще медленно (реже одного сокращения в секунду). С правой стороны сердечной трубки на желточном мешке расположен кроветворный мешочек, наполненный эритробластами. Глаза эмбрионов на этой стадии сильно пигментированы. Образовались жаберные щели. На желточном мешке вдоль тела зародыша появились первые мелкие меланофоры. Слуховая капсула заметно увеличилась. Тело эмбриона обрамлено плавниковой складкой.
Возраст 75 сут — начало кровообращения (рис. 19, в). Пигментированные меланином глаза хорошо просматриваются через оболочку икринки. Сердце активно пульсирует по замкнутой сосудистой системе. Меланофоры начинают образовываться на туловищном и хвостовом отделах эмбриона. Печень полностью обособилась от кишечной трубки и расположена на желточном мешке левее места отделения туловища. На переднем конце головного отдела образовались обонятельные ямки. Появились зачатки жаберных крышек. Железистые клетки грудных плавников сохранились только по краям их лопастей. Эмбрион активно переворачивается внутри оболочки. В связи с энергичными вращениями зародыша внутри оболочки значение гидростатической функции этого синуса утрачивается, поэтому он постепенно редуцируется.

Рисунок 19. Образование системы кровообращения (начало пигментации глаз): а — начало пигментации глаз (55 сут); б — появление форменных элементов крови (65–70 сут); в — появление системы кровообращения (75 сут)
Возраст 80–85 сут — образование разветвленной подкишечно-желточной системы кровообращения. (рис. 20, а).
На дне ротовой воронки сформировался зачаток нижней челюсти. Сосудистая сеть, покрывающая заднюю половину левой части желточного мешка, выполняет функцию эмбрионального органа дыхания зародыша.
Из головного отдела кровь течет по парным передним кардинальным венам, которые, соединяясь с разветвленными участками задней кардинальной вены в грудной области, образуют короткие кювьеровы протоки, впадающие в венозный синус сердца. В слуховой капсуле образуются полукружные каналы. Меланофоры расположены вдоль спинной части туловища, кишечной трубки и задней половины желточного мешка, омываемой кровью.
Возраст 90 сут. — исчезновение перибластического синуса и кроветворного мешочка (рис. 20, б). В головном отделе появились меланофоры. Голова отделяется от желточного мешка. Позади кишечной трубки, в области анального отверстия, образовался зачаток мочевого пузыря. Железы вылупления большими скоплениями расположены на перикардии, верхней и нижней челюстях, зачатке жаберной крышки, а также между глазами и слуховыми капсулами.
Возраст 110 сут — появление гиоидных дуг аорты (рис. 20, в). Жаберная крышка покрывает первые две жаберные дуги. Глаза покрыты серебристым пигментом — гуанином. Слуховая капсула увеличилась. Меланофоры расположены по всей поверхности желточного мешка. Печень заметно увеличивается. В области 6-го миотома появился гломус — предпочка.

Рисунок 20. Образование разветвленной подкишечно-желточной системы кровообращения: а — четверть поверхности желточного мешка покрыта сосудистой системой желточной вены (80–85 сут); б — третья часть желточного мешка покрыта сосудами желточной вены (90 сут); в — появление гиоидных дуг аорты (110 сут)
VIII этап — подвижное состояние челюстей.
Возраст 130 сут — начало движения нижней челюсти. (рис. 21, а). В их основании образовался зачаток плечевого пояса — клейтрум. Эмбрион активно шевелит грудными плавниками, создавая сильный ток жидкости под оболочкой. Нижняя челюсть подвижна и густо покрыта железами вылупления. Диаметр печени увеличился. В просвете кишечной трубки появилась складчатость.
Возраст 180 сут — появление жаберных лепестков и образование псевдобранхий (рис. 21, б). У заднего края глаз сформировался двухлопастный зачаток псевдобранхий. На жаберных дужках появились зачатки жаберных лепестков. Кишечная трубка покрыта крупными звездчатыми меланофорами, образующими полосу, которая выходит на желточный мешок. Сеть кровеносных сосудов на желточном мешке сократилась.
В природных условиях на этом этапе развития начинается вылупление и скат свободных эмбрионов с нерестилищ. На рыбоводном заводе в это время вылуплялись только ненормально развитые зародыши. У большинства из них аномалии в кровообращении. В основном преждевременно вылупляются слабые, нежизнеспособные эмбрионы, которые малоподвижны внутри оболочки. Из-за слабой подвижности эмбрионов фермент, выделяемый железами вылупления, растворяет оболочку в области головы, и эмбрион вылупляется головой вперед. Таким образом, головной отдел зародыша защемляется оболочкой икры в области сердца и погибает. Следовательно, вылупление эмбрионов из оболочки головой вперед является признаком аномального развития.
Нормально развитые эмбрионы вылупляются вперед хвостом, резким движением которого они прорывают оболочку и сбрасывают ее с себя через голову. Оболочка к концу этого этапа под действием фермента вылупления становится менее прочной. Такие икринки имеют эллиптическую форму. Кроме того, у конца загнутой к верху хорды, в нижней части хвостовой лопасти сформированы жесткие хрящевые пластины — зачатки опорных элементов костных лучей, способствующие быстрому освобождению эмбриона от оболочки.
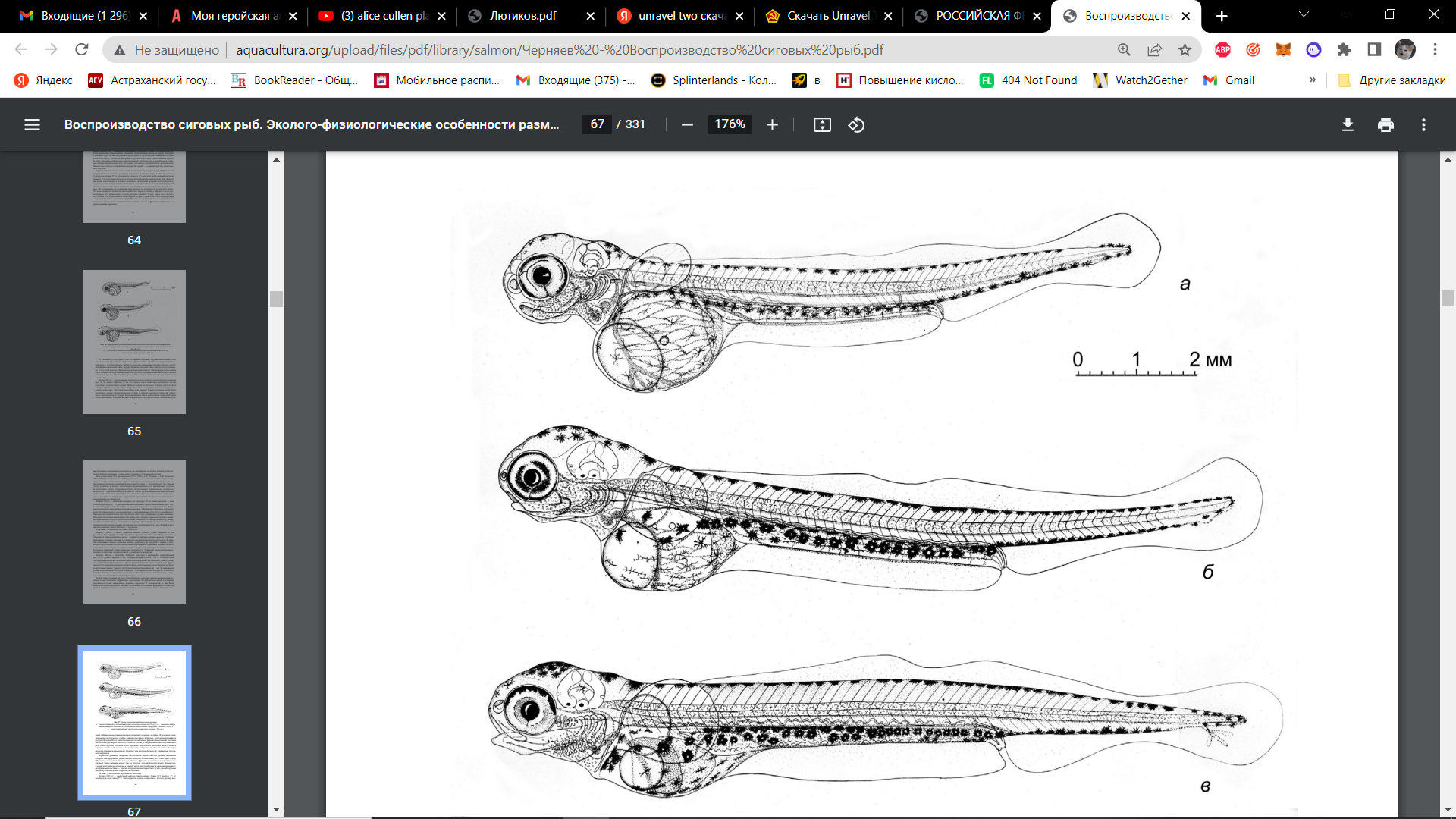
Рисунок 21. Этапы подготовки эмбрионов к вылуплению: а — начало подвижного состояния жаберно-челюстного аппарата (130 сут.); б — появление и образование жаберных лепестков и псевдобранхий (вылупление в природных условиях, 180 сут.); в — свободный эмбрион (вылупление в заводских условиях, 220 сут.)
IX этап — вылупление зародыша из оболочки.
Возраст 220 сут — свободный эмбрион (предличинка). (рис. 27, в). Запасы желтка сильно сократились, поэтому размер желточного мешка значительно уменьшился. В нем находится довольно крупная жировая капля, диаметром 0,6 мм. Печень оказывается позади желточного мешка. В кишечной трубке заметны перистальтические движения. Гломус имеет железистое строение. Псевдобранхия хорошо сформирована и омывается кровью, поступающей по гиоидной и мандибулярной дугам аорты для дополнительного обогащения кислородом поступающей в глаза крови. После вылупления зрительный анализатор начинает играть первостепенную роль при переходе личинки на внешнее питание. Зачатки жаберных лепестков еще не обеспечиваются кровью, и жаберное дыхание еще отсутствует. Дыхательная сеть кровеносных сосудов на желточном мешке почти полностью редуцирована. Хвостовая вена образует разветвления в нижней лопасти хвостовой плавниковой складки. Эта капиллярная сеть сосудов выполняет дыхательную функцию, компенсируя утрату этой роли желточным кровообращением. Жаберный аппарат, развиваясь все более, постепенно начинает выполнять функцию дыхания.
Вылупившиеся эмбрионы всплывают к поверхности воды. Они обладают положительным фототаксисом и стремятся плыть против течения. Эмбрионы держатся большими скоплениями в затишных участках. Окраска их светлеет, и они становятся трудно заметными в воде. [43]