

Сравнительная зоология позвоночных / Короткова А.А. Сравнительная анатомия беспозвоночных животных
.pdfнекоторых представителей заметны границы сегментов головы. Туловищный отдел состоял из большого количества гомономных сегментов. Анальная лопасть у большинства представителей сливалась с несколькими последними сегментами и образовывала пигидий. Таким образом, им была присуща билатеральная симметрия и гомономная сегментация.
Рис. 30. Строение трилобитов: А – с брюшной стороны, Б – со спинной стороны. 1 – головной щит, 2 –сегменты головы, 3 – сложные глаза, 4 – сегменты туловища, 5 – пигидий, 6 – антеннулы, 7 – ноги,
8 – ротовое отверстие
У ракообразных Crustacea (рис. 31) тело состоит из трех отделов – головы, груди и брюшка, каждый из которых образован сходными по строению и функциям сегментами. Голова образована акроном и четырьмя сегментами и, соответственно, несет придатки акрона – антенуллы – и четыре пары конечностей. У некоторых ракообразных с акроном сливается только сегмент антенн. Это образование носит название протоцефалона (первичной головы). В некоторых случаях сливаются воедино челюстные сегменты с образованием гнатоцефалона (челюстного отдела). Если гнатоцефалон срастается с некоторым числом грудных сегментов,
31

образуется гнатоторакс (челюстегрудь). Грудные сегменты могут сливаться с головой, образуя головогрудь. Брюшные сегменты низших ракообразных лишены конечностей, но у высших ракообразных конечности на брюшке присутствуют.
2
1 |
3 |
4
Рис. 31. Ракообразные: 1 – речной рак, 2 – жаброног, 3 – креветка, 4 – краб
Тело паукообразных (Arachnida) подразделяется на два отдела - головогрудь (просома) и брюшко (опистосома).
Головогрудь образована слиянием головы (акрона и 4 сегментов) с двумя грудными сегментами. Брюшко может быть разделено на переднебрюшие и заднебрюшие, как, например, у скорпионов. В пределах
32

класса происходит слияние сегментов брюшка, вплоть до их полного исчезновения (рис. 32). На конце тела располагается тельсон.
1 |
2 |
3
Рис. 32. Паукообразные: 1 – скорпион (1 – просома, 2 – переднебрюшие, 3 – заднебрюшие), 2 – паук птицеяд, 3 – иксодовый клещ)
Тело многоножек (Myriapoda) (рис. 33) состоит из слитной головы и туловищного отдела. Голова образована слиянием акрона и четырех сегментов, между которыми отсутствуют какие-либо видимые границы.
33

Туловищный отдел сегментирован. Число сегментов варьирует в значительной мере в пределах группы (от 10 до 170 и более). Общая гомономность метамерии у некоторых представителей нарушается следующими событиями. Так, у двупарноногих (рис. 33-3) выделяются 4 грудных сегмента, один из которых вовсе не несет конечностей, а три следующих несут по одной паре конечностей. Остальные сегменты несут по две паре конечностей, вполне гомономны и именуются брюшными. Это так называемая региональная дифференциация сегментов. У губоногих (рис. 33– 2) возможна ритмичная дифференциация симметрии, вызванная правильным чередованием сегментов разного размера.
1 |
2 |
3 |
Рис. 33. Многоножки (1 – сколопендрелла, 2 – костянка, 3 – кивсяк)
Насекомые (Insecta) (рис. 34) имеют три отдела тела – голову, грудь и брюшко, в состав которых входит достаточно стабильное количество сегментов. Так, слитная голова образована акроном и четырьмя сегментами. Грудной отдел всегда состоит из трех сегментов – переднегру-
34

ди, среднегруди и заднегруди. В состав брюшного отдела входят от 4 до 11 сегментов.
Рис. 34. Внешнее строение насекомых (черный таракан): 1 – голова; 2 – грудь; 3 – усики; 4 – конечности; 5 – крылья; 6 – брюшко
Таким образом, у насекомых выражена типичная гетерономная метамерия.
Моллюски (Mollusca) (рис. 35), как типичные билатерии, имеют двустороннюю симметрию тела. Метамерность проявляется у полиплакофор (рис. 35-1) в строении раковины, образованной 8 пластинками, и в строении мускулатуры раковины. Последняя метамерна и у моноплакофор (рис. 35-2). Помимо этого, отмечается метамерия жабр (полиплакофоры, моноплакофоры) и почек (моноплакофоры).
Двустворчатые моллюски (рис. 35-3) и головоногие моллюски (рис. 35-5) типично билатерально симметричные животные.
Брюхоногие (рис. 35-4) в своем строении сочетают билатеральную симметрию и диссимметрию. Строение головы и ноги их вполне соответствует билатеральной симметрии. Диссимметрия проявляется в спиральном строении раковины и туловищного мешка, в исчезновении парности почек, жабр, предсердий, а так же в строении нервной системы (хиастоневрия у двупредсерных и однопредсердных переднежаберных моллюсков).
35

1 |
2 |
3 |
4 |
5
Рис. 35. Моллюски: 1 – хитон, 2 – моноплакофора, 3 – перловица, 4 – виноградная улитка, 5 – кальмар
Предком современных брюхоногих моллюсков были плавающие билатерально симметричные формы. В дальнейшем эволюция этих животных шла по пути увеличения размеров раковины, в которую должно
36

втягиваться все тело животного. Первичная конусовидная раковина, увеличиваясь в размерах, стала закручиваться в спираль, что является более экономичной формой. Завиток раковины был направлен вперед. С переходом к ползающему способу передвижения завиток переместился назад, т.к., находясь в переднем положении, он давил на голову. Такое филогенетическое перекручивание (торсия) подтверждается наличием физиологической торсии в онтогенезе современных моллюсков (рис. 36). Именно этот процесс привел к появлению хиастоневрии.
Рис. 36. Поворот раковины в эволюции брюхоногих моллюсков:
1 – предковая форма, 2,3 – этапы поворота раковины завитком назад
На смену раковине, спирально закрученной в одной плоскости, приходит турбоспиральная раковина, что является более адаптивным признаком (наиболее выгодная в пространственном отношении форма). Изменяется положение раковины в связи со смещением центра тяжести
37
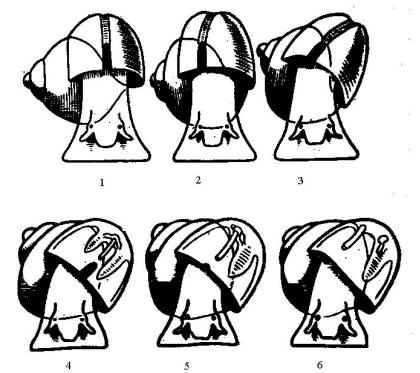
(рис. 37-1). Раковина приобретает наклон влево. Именно это влечет за собой редукцию правой части органов мантийного комплекса и появление диссимметрии (рис. 37-2).
Рис. 37. Происхождение диссимметрии брюхоногих моллюсков: 1–3 – изменение положения турбоспиральной раковины; 4–6 – развитие
диссимметрии органов мантийного комплекса
Погонофоры (Pogonophora) (рис. 38, 39) в своем строении сочетают билатерльную симметрию и гетерономную метамерию. На переднем отделе тела расположена головная лопасть и щупальца. На втором отделе расположена уздечка в виде валиков с кутикулярными килями. Следующий отдел самый длинный и называется туловищным. На конце тела расположен задний отдел. На туловище более или менее метамерно рас-
38

положены хитиноидные пластинки, лежащие на небольших возвышениях – папиллах. Они служат для закрепления животного в трубке.
Рис. 38. Погонофоры
Рис. 39. Внешнее строение погонофор (1 – головная лопасть, 2 – уздечка, 3, 10 – брюшные ресничные полоски, 4 – пластинки, 5 – задний отдел тела, задний конец третьего отдела тела, 7 – спинная папилла, 8 – железистный щиток, 9 – пояски, 11 – крупные папиллы, 12 – парные метамерные папиллы, 13 – второй отдел тела, 14 – щупальца, 15 – пиннулы.
39

Для взрослых форм иглокожих (Echinodermata) (рис. 40) характерна радиальная симметрия. Количество плоскостей симметрии зависит от количества радиусов тела животного и может быть от пяти и более. Ранняя стадия личинки – диплеврула – одинакова для всех иглокожих и имеет билатеральную симметрию. Исходя из этого обычно делают вывод, что предки иглокожих также были билатерально симметричными организмами. Таким образом, радиальная симметрия современных форм является вторичной и связана с переходом к малоподвижному образу жизни. Однако радиальная симметрия иглокожих в некоторых случаях нарушается. Так, например, у некоторых морских ежей и морских звезд наблюдается эксцентричное положение ануса, а у некоторых морских ежей и у всех голотурий проявляется двусторонняя симметрия, являющаяся в данном случае третичной. В строении же скелета и амбулакральной системы отчетливо проявляется метамерность.
1 |
2 |
3
Рис. 40. Иглокожие: 1 – морская звезда, 2 – морской еж, 3 – офиуры
40