

Сравнительная зоология позвоночных / Короткова А.А. Сравнительная анатомия беспозвоночных животных
.pdfраспада, или схизоцелем. Присутствует у круглых червей. Схизоцель не имеет собственных стенок, заполнен полостной жидкостью, которая выполняет опорную и распределительную функции. В некоторых случаях именно схизоцель называют первичной полостью тела. Однако более справедливо называть так полость бластулы – бластоцель, поскольку именно он появляется в онтогенезе первым в качестве полости. При гаструляции бластоцель исчезает и никаких производных не дает, т.е. является провизорным органом.
Целом, или вторичная полость, имеет собственный целомический эпителий и выводные протоки – целомодукты. Таким образом, целом представляет собой самостоятельный орган. Целом присущ в той или иной мере всем трохофорным животным, а также вторичноротым. Происхождение целома представляет значительную проблему, единого взгляда на которую нет (Беклемишев В.Н., 1964). Существует несколько более или менее правдоподобных гипотез. Наиболее приемлемой является энтероцельная гипотеза И.И. Мечникова, в соответствии с которой целом является гомологом периферического отдела гастроваскулярной системы кишечнополостных.
Образование целома в онтогенезе у первичноротых и вторичноротых происходит различным образом. У первичноротых на стадии бластулы выделается один бластомер, который делится на две клетки – телобласты (рис. 93).
А |
Б |
В |
Рис. 93. Схема развития целома полихет: А, Б, В – поперечные срезы на последовательных стадиях развития червя, 1 – эктодерма,
2 – кишечник (энтодерма), 3 – мезодермальные полоски, 4 – целомический эпителий, 5 – целом, 6 – спинная и брюшная брыжейки, 7 – спинной кровеносный сосуд, 8 – брюшной кровеносный сосуд, 9 – мышцы,
10 – брюшная нервная цепочка
91
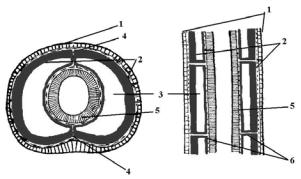
При гаструляции телобласты располагаются по бокам от бластопора В последствии они дают мезодермальные полоски, из которых формируются клетки целомического эпителия. Этот способ образования мезодермы носит название телобластического. Мезодермальные полоски под влиянием внешней сегментации так же сегментируются, а затем в них образуются полости – целом.
У вторичноротых мезодерма образуется энтероцельно, путем отшнуровывания от энтодермальной первичной кишки. Данные сравнительной анатомии и сравнительной эмбриологии говорят в пользу независимого происхождения целома у первичноротых и вторичноротых и таким образом, их целом негомологичен (Беклемишев В.Н., 1964).
Из первичноротых наиболее примитивный целом у олигомерных кольчатых червей. Он представлен исключительно половыми полостями, с собственными протоками – целомодуктами, и присутствует в отдельных постларвальных сегментах. В остальных сегментах между внутренними органами располагается мезенхима с обширными схизоцельными пространствами. У полимерных аннелид целом построен по единому плану. В головной и анальной лопастях отсутствует. В ларвальных сегментах возникает одновременно. В постларвальных сегментах образуется постепенно в связи с образованием сегментов. Целомические мешочки в каждом сегменте парные (рис. 94).
А |
Б |
Рис. 94. Схема поперечного (А) и продольного (Б) разреза тела кольчатого червя: 1 – кожно-мускульный мешок, 2 – целомический эпителий, 3 – целом, 4 – мезентерий, 5 – кишка, 6 – диссепименты
92
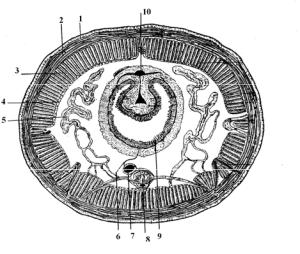
Так, у дождевых червей редуцирован спинной мезентерий, в результате чего в каждом сегменте формируется единый целомический мешок (рис. 95). Целом выполняет выделительную, опорную и половую функции.
Рис. 95. Поперечный разрез тела дождевого червя: 1 – эпителий, 2 – кольцевые мышцы, 3 – продольные мышцы, 4 – целомический эпителий, 5 – метанефридий, 6 – мезентерий, 7 – брюшной кровеносный сосуд, 8 – брюшная нервная цепочка, 9 – кишечник, 10 – спинной кровеносный сосуд
Целом пиявок претерпевает наиболее значительные изменения.
Уэтих животных целом вытесняется паренхимой, которая заполняет пространства между внутренними органами (рис. 96). Целом редуцирован до системы каналов и, в некоторых случаях, принимает на себя функцию кровеносной системы. Причем степень редукции целома различна у пиявок разных групп.
Ущетинконосных пиявок вторичная полость тела хороша развита. При этом полностью сохраняется кровеносная система (рис. 96 – А).
Ухоботных пиявок целом сохраняется в виде системы лакун, среди которых наиболее развиты продольные спинная, брюшная и боковые лакуны. Эти лакуны сообщаются между собой более мелкими лакунами. В спинной и брюшной лакунах залегают кровеносные сосуды (рис. 96 – Б). У челюстных пиявок редукция целома и разрастание заходят так же далеко, как и у хоботных. Однако в этом случае кровеносная система редуцируется полностью и ее функции выполняет система целомических лакун
(рис. 96 – В).
93
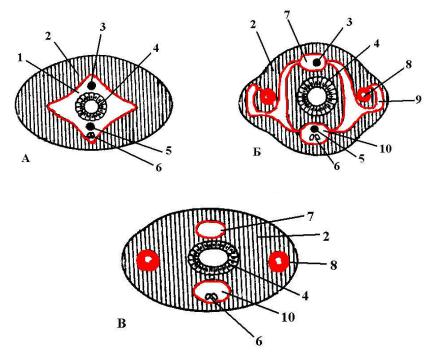
Рис. 96. Изменение целома у различных пиявок (схема поперечного среза):
А– щетинконосная пиявка, Б – хоботная пиявка, В – челюстная пиявка, 1 – целом, 2 – паренхима, 3 – спинной кровеносный сосуд, 4 – кишка,
5 – брюшной кровеносный сосуд, 6 – брюшная нервная цепочка, 7 – спинная лакуна целома, 8 – боковая лакуна целома, 9 – пузырь боковой
лакуны целома, 10 – брюшная лакуна целома
У моллюсков сильно развита паренхима, заполняющая пространство между внутренними органами. Целом обычно присутствует в виде двух полостей – полости гонады и полости околосердечной сумки (перикардия). Каждый из отделов целома имеет свои протоки. При этом целомодукты полового участка целома функционируют соответственно как половые протоки, а целомодукты перикардия – как почки. У головоногих половой и перикардиальный отделы целома сообщаются. У наутилуса оба отдела очень обширны, половой отдел образует настоящую полость тела. У осьминогов полость тела редуцирована и сведена до системы узких, тонкостенных каналов, соединяющих полость гонады с почкой и
94
внешней средой. Таким образом, целом редуцирован и специализирован, но связь между двумя отделами сохраняется. У остальных моллюсков эти отделы не соединены.
Весьма своеобразна судьба целома у членистоногих. При эмбриональном развитии образуются целомические мешочки. Они располагаются метамерно в ларвальных и постларвальных сегментах, а также в голове. Затем они теряют эпителиальную поляризацию и дают начало аморфной ткани – целенхиме. Целенхима дает начало мышцам, выполняющей (соединительной) ткани, жировому телу. Поскольку стенки целома распадаются целомические полости сливаются с сохранившимся схизоцельным пространством. Образующаяся полость тела называется смешанной,
или миксоцелем.
Большинство ракообразных утратили целом даже на эмбриональных стадиях. У некоторых он сохраняется в виде провизорного органа. В одельных случаях целом дает производные, в частности стенки гонад и антеннальные железы. Целом многоножек сохраняется на эбриональных стадиях, позднее дает гонады. Таким образом полость гонад представляет собой целомическую полость.
Среди насекомых степень редукции целома напрямую связан с уровнем организации. У низших насекомых целом достаточно отчетливо выражен на эмбриональных стадиях. У насекомых с полным метаморфозом зародышевый целом претерпевает значительную редукцию, а у некоторых групп двукрылых отсутствует вовсе.
Паукообразные по степени сохранности зародышевого целома занимают первое место среди членистоногих. За счет него образуются гонады и коксальные железы.
Вторичноротые беспозвоночные животные имеют хорошо развитый целом. У погонофор в первом сегменте присутствует непарный целомический мешок, а в трех других – парные целомические мешки. Целомические каналы заходят в щупальца. Производными целома являются гонады.
У иглокожих целом развит наиболее сильно и дает серию производных. На стадии ранней личинки диплеврулы образуются парные целомические мешки, которые впоследствии делятся на три парные части (рис. 97). Из задних зачатков развивается собственно целом. Его эпителий дает начало мышцам, скелету, соединительной ткани, кровеносной системе, гонадам (рис. 98 – 17, 18). Целом парный (рис. 98 – 5) и заполнен целомической жидкостью. Правый передний и средний зачатки редуцируются.
Левый передний зачаток трансформируется в каменистый канал, мадрепоровую пластинку, осевой орган (рис. 98 – 4, 15), осевые синусы (рис. 98 – 3, 14) и псевдогемальные каналы.
95
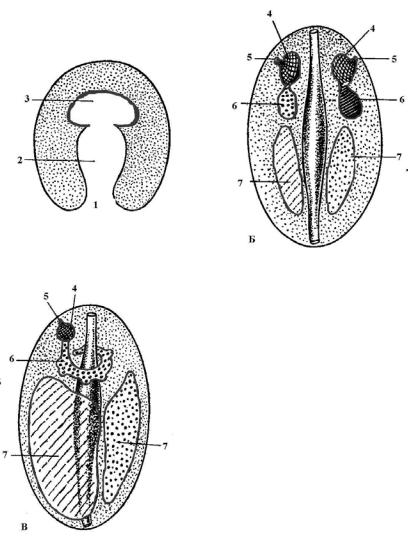
А
Рис. 97. Развитие целома у иглокожих: А – образование первичного целомического мешка, Б – дифференцировка трех пар целомических мешков, В – дальнейшее изменение целомов, 1 – бластопор, 2 – гастральная полость, 3 – первичный целомический мешок, 4 – левый и правый передние целомы, 5 – гидропоры, 6 – левый и правый средние целомы, 7 – левый и правый задние целомы
96
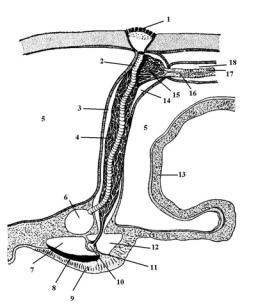
Из левого среднего зачатка целома образуются остальные части амбулакральной системы. За счет стенок целома образуются гипоневральный и эктоневральный отделы нервной системы (рис. 98 – 8, 9). Этот факт образования нервной системы из периферического фагоцитобласта является почти единственным исключением во всем животном мире. Напомним, что в остальных случаях нервная система является производным кинобласта.
Рис. 98. Схема осевого комплекса морской звезды:1– мадрепоровая пластинка, 2 – каменистый канал, 3 – левый осевой синус, 4 – оральный отдел осевого органа, 5 – целом, 6 – кольцевой амбулакральный сосуд, 7 – наружное псевдогемальное кольцо, 8 – гипоневральный отдел нервной системы, 9 – эктоневральный отдел нервной системы, 10 – кольцевой
оральный кровеносный сосуд, 11 – септа, 12 – внутреннее псевдогемальное кольцо, 13 - стенка желудка, 14 – правый осевой синус, 15 – аборальный отдел осевого органа, 16 - аборальный кровеносный сосуд, 17 – половой столон, 18 – половой синус
Амбулакральная система (рис. 98) образована мадрепоровой пластинкой, каменистым каналом, кольцевым каналом и радиальными кана-
97
лами. Мадрепоровая пластинка расположена на аборальной стороне и прободена порами. Каменисный канал залегает внутри осевого органа. На кольцевом канале располагаются полиевы пузыри, которые являются резервуарами жидкости. На радиальных каналах расположены ампулы и амбулакральные ножки. Эта система заполнена жидкостью и выполняется функцию движения.
Псевдогемальная система также представлена комплексом каналов, расположенных к периферии от амбулакральной системы (рис. 98 – 7, 12). Жидкость псевдогемальной системы сходна с полостной жидкостью. Смыкаясь, стенки псевдогемальных каналов образуют септы (рис. 98 – 11), в которых залегают лакуны кровеносной системы (рис. 98 – 10, 16). Псевдогемальная система участвует в трофической функции.
Вопросы и задания для самоконтроля. 1. Каково происхождение и функции паренхимы? 2. Для каких животных характерно наличие паренхимы? 3. Что такое схизоцель? 4. Каково происхождение схизоцеля? 5. Для каких животных характерно наличие схизоцеля? 6. Что такое целом? 7. Каково происхождение целома? 8. Как происходит образование целома в онтогенезе? 9. Каковы особенности целома кольчатых червей и моллюсков? 10. Какова судьба целома у членистоногих? 11. Что такое миксоцель? 12. Каково строение целомаиглокожих? 13. Каково строение целомапогонофор?
4.3. Сократительно-двигательный аппарат
Движение является одним из важных признаков животного организма. Первоначально она трансформируется из функции захвата пищи.
Простейшие двигаются за счет перетекания недифференцированной протоплазмы (амебы), жгутиков (жгутиковые), ресничек (инфузории), мионем. Последние представляют собой сократимые волокна, расположенные в цитоплазме. Встречаются у грегарин и инфузорий.
У большинства многоклеточных также имеются сократимые волокна, которые в принципе соответствуют мионемам простейших. Совокупность сократимых элементов тела представляет мышечную систему животного. Она входит в состав сократительно-двигательного аппарата. Помимо этого он включает в себя опорные элементы (опорные пластинки, различного рода скелеты). Последние являются антогонистами сократимых элементов, а также объединяют их в единое целое. Уровень развития сократи- тельно-двигательного аппарата зависит от уровня организации животного,
98
в свою очередь, оказывая на него большое влияние (В.Н. Беклеми-
шев, 1964).
Губки, как наиболее примитивные многоклеточные, не имеют сократимых элементов. Вся двигательная активность их сводится к замыканию пор за счет движения протоплазмы пороцитов, а также к медленному закрыванию оскулума. Однако подавляющее большинство губок имеет скелет, состоящий из известковых, кремневых или спонгиновых игл. Они вырабатываются специализированными клетками: в первых двух случаях – склеробластами, в последнем – спонгиобластами. Существует также вариант кремнерогового скелета, формирующегося за счет деятельности обоих видов клеток. Склетные образования губок выполняют опорную функцию.
Движение кишечнополостных осуществляется за счет сокращения миофибрилл эпителиально-мускульных клеток. Как отмечалось ранее, они располагаются как в эктодерме, так и в энтодерме. Опорную функцию выполняет бесструктурная мезоглея, лежащая между упомянутыми слоями клеток. Наиболее просты движения полипов. Однако у коралловых полипов уже происходит дифференциация продольных и кольцевых мышечных волокон в эктодерме, что способствует более разнообразным движениям. В связи с этим мезоглея сильно развита и имеет волокнистую структуру.
Дифференцировка мышечных волокон наблюдается и у медуз. Причем все они лежат на нижней стороне зонтика (радиально и по краю) и сжимают его при движении. Обратное расправление тела обеспечивается упругим сопротивлением сильно развитой студенистой мезоглеи. Таким образом обеспечивается реактивное движение.
У гребневиков впервые формируется хорошо обособленная мышечная ткань, лежащая в толще мезоглеи. Последняя по обыкновению выполняет скелетную функцию. Различаются кольцевые и продольные мышечные волокна под эктодермой, вокруг кишечника. Радиальные мышцы идут от кишечника к стенкам тела. Достаточно сильно развиты мускулы щупалец. Иногда присутствуют сфинктеры вокруг рта. Однако, как известно, гребневики двигаются за счет гребных пластинок (суть – сохраняют мер- цательно-локомоторную функцию кинобласта пожизненно). Таким образом вся мускулатура в основном обеспечивает хватательные и глотательные движения, что вполне соответствует хищному образу жизни этих животных. У плоских червей наличествует хорошо развитый кожномускульный мешок, образованный эпителием и несколькими слоями мышц. Эти слои имеют кинобластическое происхождение. Для турбеллярий характерно три слоя мышц – кольцевые, косые и продольные (рис. 99).
99

Рис. 99. Схема строения кожномускульного мешка и паренхимы турбеллярии: 1 – ресничный эпителий, 2 – кольцевые мышцы, 3 – косые мышцы, 4 – продольные мышцы, 5 – дорзо-вентральные мышцы, 6 – паренхима
Рис. 100. Схема строения кожномускульного мешка дигенетического сосальщика: 1 – тегумент, 2 – базальная мембрана, 3 – кольцевые мышцы, 4 – продольные мышцы, 5 – паренхима, 6 – погруженная часть эпителия
100