

Сравнительная зоология позвоночных / Короткова А.А. Сравнительная анатомия беспозвоночных животных
.pdfИх наличие обусловлено сплющенностью глотки (глоточная плоскость) и расположением щупалец (щупальцевая плоскость). Таким образом, формируется двулучевая симметрия. Однако этот тип симметрии не охватывает организацию гребневиков полностью.
Расположение некоторых органов формирует иные плоскости симметрии. Так, пружинки статолита и четыре отходящие от них мерцательные полоски, обуславливают неполную четырехлучевую симметрию, а 8 рядов гребных пластинок и 8 меридиональных каналов создают неполную восьмилучевую симметрию (рис. 21).
Рис. 21. Аборальный орган: 1 – ресничный купол, 2 – статолит, 3 – пружинящие дужки, 4 – мерцательные бороздки, 5 – поры купола.
Следующим этапом развития является формирование билатеральной симметрии. В настоящее время существует множество взглядов на происхождение билатеральной симметрии и билатерально симметричных животных. Классические гипотезы по этой проблеме можно объединить в несколько групп (Малахов, 2004).
Наибольшее распространение получили планулоиднотурбеллярные гипотезы, сторонниками которых были, в частности, В. Н. Беклемишев, А. В. Иванов и Ю. В. Мамкаев. Согласно этим воззрениям предками Bilateria были организмы, напоминавшие паренхимул или планул (рис. 22).
На одном полюсе этих организмов располагался чувствительный аборальный орган, а на другом – рот. Они плавали в толще воды при помощи ресничек, а затем перешли к донному образу жизни. При этом им было свойственно активное поступательное движение, что, как и в случае с простейшими, привело к появлению билатеральной симметрии. Первичные Bilateria не имели сквозного кишечника и целома. В современной фауне к ним наиболее близки бескишечные турбеллярии. Все упомянутые ученые считали, что радиально-симметричный предок стал ползать
21
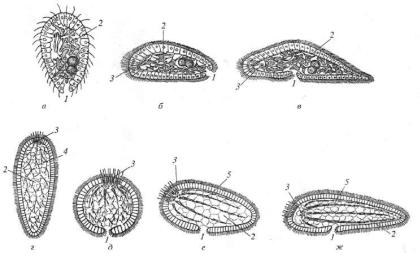
по дну на боковой поверхности (точнее – на одной из антимер), которая и стала брюшной стороной. Рот при этом располагался на заднем конце тела.
Рис. 22. Происхождение Bilateria согласно планулоидно-турбеллярным гипотезам: а – в – стадии оседания радиально-симметричного предка на антимеру, которая становится брюшной стороной Bilateria (по Иванову, 1968); г–ж – оседание радиально-симметричного предка на оральный полюс, который становится брюшной стороной (по Hyman, 1951). Обозначения: 1 — рот, 2 – паренхима, 3 – аборальный нервный центр, 4 – нервное сплетение, 5 – нервные стволы.
Для трохофорных животных (аннелид, членистоногих, моллюсков и др.) В. Н. Беклемишев (1944, 1964) предполагал другой путь происхождения от радиально-симметричных предков. Он считал, что их предками были организмы, сходные с геккелевской гастреей. При этом гастрея, опустившись на дно, стала ползать на оральной, а не на боковой стороне. Предками же вторичноротых В. Н. Беклемишев (1964) считал формы, похожие на гребневиков и перешедшие на одной из антимер, лежащей в глоточной плоскости.
Сторонники архицеломатных гипотез предполагали, что предками Bilateria были четырехлучевые полипы, гастральная полость которых была разделена перегородками на четыре камеры. Они стали ползать на
22
ротовой поверхности, которая превратилась в брюшную сторону. Их первичный рот превратился в щелевидный, сомкнулся посередине, а образовавшиеся отверстия стали ртом и анусом. Камеры гастральной полости, отделившись, дали начало целомическим мешкам.
Существует также гипотеза колониального происхождения Bilateria, появившаяся в 2000 году и названная В. В. Малаховым (2004) «остроумной». В ней индивидуум Bilateria рассматривается как результат интеграции целой колонии особей кишечнополостных. При этом метамерно расположенные полипы колонии превращаются в отдельные органы билатерально-симметричного индивидуума (например, в метамерные гонады). Эти соображения не согласуются с данными эмбрионального развития Bilateria, в онтогенезе которых нет никаких следов слияния нескольких особей.
На основании анализа формирования билатеральной симметрии в эмбриогенезе и новейших палеонтологических данных В.В. Малаховым (2004) были высказаны новые взгляды на происхождение Bilateria. Ранние стадии развития подчиняются радиальной симметрии. Формирование билатеральной симметрии начинается у примитивных билатерий только после стадии гаструлы. Это происходит путем разрастания одного из секторов гаструлы, а именно того, который станет спинной стороной личинки. Аборальный орган смещается вперед, а бластопоральная сторона оказывается брюшной стороной личинки. Это характерно для подавляющего большинства групп Bilateria (плоских червей, аннелид, моллюсков, иглокожих). Во многих группах Bilateria бластопор вытягивается и приобретает щелевидную форму. Замыкание щелевидного бластопора приводит к формированию трубчатого кишечника. Рот и анус Bilateria есть производные переднего и заднего концов щелевидного бластопора.
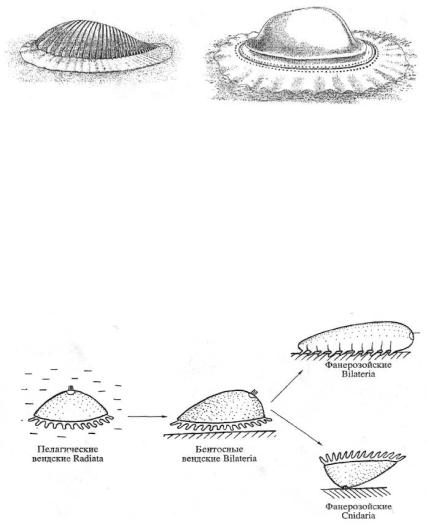
Согласно взглядам В.В. Малахова (2004), в вендском периоде существовала обширная фауна радиально-симметричных двухслойных кишечнополостных. Часть их представителей перешла к ползанию по субстрату на оральной поверхности, что привело к формированию билатеральной симметрии (рис. 23).
Вендские Bilateria, скорее всего, не были еще трехслойными животными. Их кишечная полость могла быть связана с внешней средой длинным щелевидным ртом, тянущимся вдоль брюшной (оральной) стороны. Возможно, что метамерно расположенные карманы гастральной полости тоже еще не были отделены от ее центральной части. Такие би- латерально-симметричные кишечнополостные стали родоначальниками фанерозойских трехслойных животных. При этом щелевидный бластопор
23
сомкнулся посредине, а гастральные карманы отделились от центрального трубчатого кишечника.
1 2
Рис. 23. Реконструкции вендских билатерально-симметричных организ-
мов (по Федонкину, 2001): 1 – Dickinsonia costata, 2 – Kimberella quadrata
Сохранение аборального нервного центра у личинок современных Bilateria говорит о том, что их радиально-симметричные предки не были сидячими формами. Скорее всего, они вели пелагический образ жизни, и перешли к донному существованию как подвижные организмы, ползающие на оральной поверхности.
Не исключено, что вендские билатерально-симметричные кишечнополостные были предками не только трехслойных Bilateria, но и двухслойных фанерозойских кишечнополостных (рис. 24).
Рис. 24. Схема, иллюстрирующая происхождение билатерально-симметричных животных (по Малахову, 2004)
24
Среди них у коралловых полипов наблюдается билатеральная симметрия, что практически несовместимо с сидячим образом жизни. Проявление билатеральной симметрии у современных и ископаемых кораллов, скорее всего, представляет собой наследие симметрии предков, которая постепенно утрачивается в результате прикрепленного образа жизни
(Малахов В. В., 2004).
Итак, вероятные предки трехслойных Bilateria оказались весьма сложными в морфологическом отношении организмами,– обладавшими сквозным кишечником, метамерией и целомом.
Таким образом, фанерозойские кишечнополостные и трехслойные билатерально симметричные животные – это сестринские группы, потомки вендских билатерально-симметричных кишечнополостных. Гребневики лишены признаков билатеральной симметрии. Сохранение полной осевой симметрии и наличие сложного аборального органа говорит о том, что гребневики – потомки древних пелагических Eumetazoa. Современные ползающие и сидячие гребневики – это формы, вероятно, относительно недавно перешедшие к донному образу жизни. Они передвигаются на оральной стороне, выделенный передний полюс у них отсутствует и сохраняется двулучевая симметрия. Единственный сидячий гребневик Tjalfiella прикрепляется не аборальным полюсом, а оральной стороной.
Данные сравнительной молекулярной биологии указывают на то, что гребневики обособились до разделения современных кишечнополостных и трехслойных билатерий. Можно предполагать, что гребневики - это единственная группа первично радиально-симметричных организмов среди современных Eumetazoa.
Итак, перейдем к рассмотрению симметрии Bilateria. Наиболее примитивными из них являются плоские черви (Plathelminthes), а среди последних – ресничные черви, или турбеллярии (Turbellaria).
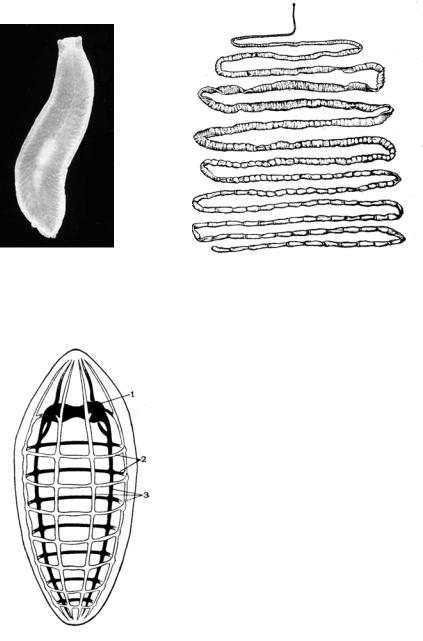
Упредставителей плоских червей (Plathelminthes) (рис. 25), помимо двусторонней симметрии, проявляется и радиальная, особенно в строении кожной мускулатуры и нервной системы (рис. 26), а также присосок дигенетических сосальщиков (Trematoda) и прикрепительного аппарата моногенетических сосальщиков (Monogenoidea).
Уленточных червей (Cestoda) лучевая симметрия заметна также в строении сколексов (рис. 27). Помимо этого, их тело делиться на отдельные сегменты (рис. 25), в которых метамерно повторяется половая система. Таким образом, билатеральная симметрия нарушается и метамерным строением тела.
25
1 |
2 |
Рис. 25. Плоские черви: 1 – планария, 2 – бычий цепень
Рис. 26. Схема строения нервной системы плоских червей (1 – мозговой ганглий, 2 – комиссуры,3 – нервные стволы)
26
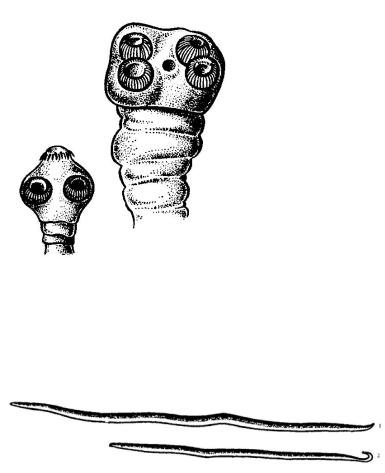
Рис. 27. Строение сколексов ленточных (1 – свиной цепень, 2 – бычий цепень)
1 |
2 |
Для круглых червей (Nemathelmintes)(рис. 28) так же характерно, помимо общей билатеральной симметрии, присутствие лучевой симметрии в строении кожно-мускульного мешка и нервной системы. Строение гонад диссимметрично.
Рис. 28. Аскарида: 1 – самка, 2 – самец
Строение следующих двух типов беспозвоночных – кольчатых червей (Annelidae) и членистоногих (Arthropoda), так же представляет собой сочетание билатеральной симметрии и метамерии.
Метамерия может рассматриваться как особый вид симметрии, впрочем, несколько условно (Беклемишев, 1964).
Метамерно-симметричным является такое тело, которое совпадает само с собой при передвижении его вдоль прямой линии – оси переноса,
27
или оси метамерии – на некоторое расстояние, обозначаемое как длина метамеры или как шаг поступательной симметрии.
Части, повторяющиеся вдоль оси называются метамерами. Тело животного при этом состоит из большого количества повторяющихся метамер, или сегментов.
Метамерность может быть гомономной или гетерономной. При гомономной метамерии все сегменты тела одинаковы. При гетерономной метамерии сегменты тела различны. Группы сходных сегментов образу-
ют отделы тела, или тагмы.
Метамерия достаточно распространена в животном мире. Она наблюдается уже начиная с простейших, в частности, в строении раковин фораминифер. Как уже говорилось, метамерия, причем гомономная, присутствует у цестод.
Тело кольчатых червей (Annelida) состоит из головной лопасти, туловищного отдела и анальной лопасти. Они сохраняют выраженную билатеральную симметрию, на которую, как уже говорилось, накладывается метамерия (рис. 29).
Головная лопасть полихет (Polychaeta) (рис. 29-1) образована простомиумом и перистомиумом (рис. 29-2). Простомиум несет пальпы, усики и глаза. На перистомиуме располагается ротовое отверстие. Перистомиум образуется слиянием двух ларвальных сегментов. В этом случае наблюдается процесс цефализации, заключающийся в образовании головной лопасти и вовлечении в ее состав туловищных сегментов, а также некоторое нарушение гомономности сегментации за счет отличий в строении сегментов головной лопасти
Метамерия туловищных сегментов гомономная, проявляется не только во внешнем строении, но и в строении и расположении внутренних органов, в частности целомических мешков. Однако у кольчатых червей сегменты тела отличаются временем и способом закладки, а также анатомическим строением, что нарушает общую гомономность сегментации. Так называемые ларвальные сегменты образуются на определенной стадии личинки. В ларвальном отделе возникает внешняя сегментация и параподии. Внутренняя сегментация протекает в зависимости от внешней путем сегментирования мезодермальных зачатков. Затем на заднем конце личинки формируется зона роста, за счет которой тело продолжает расти в длину. От оставшихся мезодермальных полосок отделяются парные мезодермальные зачатки, что определяет формирование внешней сегментации. Одновременно формируется и кожно-мускульный мешок. В результате образуются постларвальные сегменты. В ларвальнх сегментах отсутствуют половые железы и целомодукты. Брюшная нервная цепочка
28

лежит в них глубже. Имеются также отличия в строении кровеносной системы.
.
1 |
2 |
3 |
4 |
5 |
Рис. 29. Кольчатые черви: 1 – нереида, 2 – головная лопасть полихет (1 – щупальца, 2 – пальпы, 3 – простомиум, 4 – перистомиум, 5 – параподия туловищного сегмента), 3 – эпитокия у палоло,
4 – дождевой червь, 5 – пиявка.
29
Кроме того, гетерономность сегментации проявляется у полихет в случае цикла развития с эпитокией (рис. 29-3). При этом наблюдается резкая перестройка внешнего и внутреннего строения сегментов, содержащих гонады. Сегменты эпитокной части расширяются, изменяется форма параподий. До момента разделения эпитокной и атокной частей существует гетерономная метамерия.
Гетерономность присутствует у сидячих полихет. Возможна даже дифференцировка тела на более мускулистый грудной и брюшной отделы. Последний выполняет обычно половые функции.
Вцелом, гетерономность появляется в разных группах независимо
ине является превалирующим направлением изменения плана строения полихет.
Уолигохет (Olygochaeta) (рис. 29-4) достаточно четко выражено деление на ларвальные и постларвальные сегменты, различия между которыми в основном проявляются во внутреннем строении. Внешне сохраняется гомономная метамерия. Количество сегментов обычно велико
иможет доходить до 600.
Упиявок (Hirudinea) (рис. 29-5) число сегментов обычно небольшое. За счет головной лопасти и трех сегментов образуется передняя присоска, а за счет семи последних сегментов – задняя. Наличие присосок нарушает общую гомономность метамерии.
По плану строения членистоногие (Arthropoda) не отличаются от кольчатых червей и также имеют акрон, ряд метамер, анальную лопасть - тельсон. Метамерия туловища проявляется во внешней сегментации, расположении придатков и нервного ствола, скелета, мускулатуры, кровеносной системы. Гомономная метамерия наблюдается у трилобитов, листоногих раков и многоножек. У остальных членистоногих сегменты морфофункционально различаются и образуют тагмы. При этом соответственно, формируется гетерономная метамерия. Наиболее выражена гетерономность у хелицеровых. Обычно выделяют три тагмы (отдела тела)
–голову, грудь и брюшко. Голова состоит из акрона и четырех сегментов. Количество сегментов грудного и брюшного отделов сильно варьирует в пределах типа.
Усложнение метамерии от трилобитов к высшим хелицеровым проявляется в дифференциации трех отделов тела и конечностей, фиксации количества сегментов в каждой тагме, редукции части сегментов, а иногда – в интеграции тела и исчезновении сегментации.
Тело трилобитов (Trilobita (рис. 30) подразделялось на голову, туловищный отдел и анальную лопасть. Голова образовалась путем слияния акрона и четырех послеротовых сегментов. На спинной стороне у
30