

metodichka_Lipidy_2020
.pdfВ то же время в области повреждения сосудистой стенки активируются тромбоциты, которые секретируют тромбоксаны (ТХА2). Они активируют тромбоциты и стимулируют их агрегацию, а действуя на рецепторы эндотелиальных клеток, вызывают сужение просвета сосуда в области повреждения.
Перекисное окисление липидов.
Перекисное окисление липидов (ПОЛ) представляет собой свободнорадикальный процесс, который происходит без участия ферментов.
В качестве липидов, атакуемых свободными радикалами, могут использоваться остатки высших жирных кислот, включённые в состав мембранных фосфолипидов. В бо́льшей мере подвержены воздействию свободных радикалов остатки полиненасыщенных жирных кислот (линолевой, линоленовой, арахидоновой). Реакции ПОЛ инициируются активными формами кислорода.
Подобно другим свободнорадикальным процессам ПОЛ протекает в три стадии:
•инициация свободно радикальной цепи.
•продолжение цепи (элонгация).
•обрыв цепи.

На первой стадии происходит образование свободных радикалов, которые представляют собой атомы или молекулы, включающие в свою структуру атомы, содержащие неспаренные электроны. Характерным особенностью свободных радикалов является их высокая реакционная способность. Свободные радикалы могут возникать в качестве промежуточных или побочных продуктов метаболизма в различных путях обмена веществ.
Наиболее интенсивно активные формы кислорода образуются в окислительновосстановительных системах: дыхательной цепи митохондрий, электронтранспортной цепи микросом, а также некоторых ферментативных реакциях, катализируемых флавиновыми дегидрогеназами (например, ксантиноксидазой).
К активным формам кислорода относятся:
Н2О2 – перекись водорода, О2–. – супероксид-анион, ОН. – гидроксильный радикал, которые образуются в результате последовательного одноэлектронного присоединения 4-х электронов к 1 молекуле воды в реакции:
Конечный продукт этих реакций – вода, но по ходу реакций образуются свободные радикалы – активные формы кислорода.
Из всех известных свободных радикалов, образующихся в клетках, наиболее
реакционноспособным является ОН. (гидроксильный радикал). Он может возникать при неферментативном распаде перекиси водорода с участием восстановленного железа (Fe2+) в реакции Фентона:
H2O2 + Fe2+ -> ОН. + ОН– + Fe3+

Образовавшиеся свободные радикалы, вступают во взаимодействие с липидными молекулами (LH), приводя к появлению свободных радикалов липидов
(L.): OH. + LH àL. + H2O.
Образование свободных радикалов липидов завершает первую стадию процесса перекисного окисления липидов.
На второй стадии перекисного окисления липидов, свободные радикалы липидов (L.) взаимодействуют с молекулярным кислородом (О2).
В результате этого образуются новые липидные радикалы (L.) и продукты их превращений – липоперекиси (LOOН)
L. + О2 à LOO. + LHà LOOН + L.
Возникающие липидные радикалы и перекиси липидов далее подвергаются превращениям, в результате которых происходит лавинообразное образование новых липидных радикалов.
При этом один свободный радикал, возникший на первой стадии процесса, является источником сотен и тысяч других радикалов. Лавинообразная генерация радикалов происходит без участия ферментов. За счёт неферментативного взаимодействия липидных молекул, липидных радикалов и продуктов их превращений, на стадии продолжения (элонгации) цепи в процесс радикалообразования вовлекаются всё новые и новые липидные молекулы. Некоторая часть перекисей липидов может отвлекаться от процесса радикалообразования и использоваться в различных химических реакциях, в результате которых образуются альдегиды, кетоны, спирты и эпоксиды. Все указанные продукты перекисного окисления липидов обладают высокой реакционной способностью. Их накопление приводит к повреждению клетки.

Процесс образования свободных радикалов может обрываться под влиянием различных факторов. При этом наступает заключительная стадия перекисного окисления липидов – стадия обрыва цепи. Обрыв цепи может быть связан с действием
антиоксидантов. Механизм действия антиоксидантов (АO) описывается схемой L. +
АО-Н àАO. + LH.
Антиоксиданты выступают в роли донора электрона для свободного радикала. Принимая дополнительный электрон, свободный радикал трансформируется в стабильную молекулу, а антиоксидант при этом превращается в свободный радикал.
Радикалы антиоксидантов обладают низкой реакционной способностью и поэтому не способны превращать липидные молекулы в свободные радикалы.
В живых организмах встречается большое количество веществ и, в том числе продуктов метаболизма, проявляющих свойства антиоксидантов. К ним относятся витамин Е (альфа-токоферол), мочевина, мочевая кислота, билирубин, этанол и др.
Окисление полиненасыщенных кислот
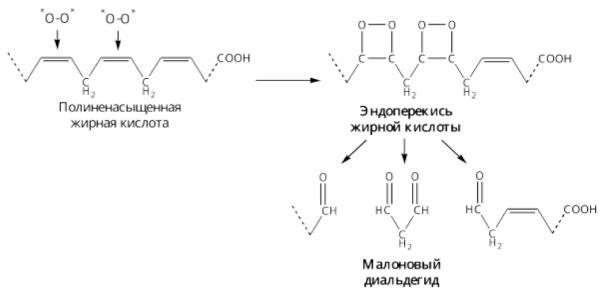
За счет образования гидроксидов жирных кислот в гидрофобном слое мембран появляются гидрофильные зоны и в клетки могут проникать: вода, ионы натрия, кальция, что приводит к набуханию клеток и их разрушению.
Процессы ПОЛ усиливаются при избытке катехоламинов (стресс), гипоксии, ишемии, снижении антиоксидантной защиты, повышенном содержании ненасыщенных кислот. Активация ПОЛ происходит при многих заболеваниях: воспалении, дистрофии, болезни Паркинсона, атеросклерозе, развитии опухолей.
Физиологическое значение реакций ПОЛ:
•регулируется проницаемость биологических мембран
•регулируют активность мембранных ферментов, реакции окислительного фосфорилирования;
•участвуют во внутриклеточном делении, передаче генетической информации
•участвуют в адаптации организма.
Повышение активности ПОЛ приводит к:
•разрушению клеточных мембран, повреждению и гибели клеток
•модификации ЛП, особенно ЛПНП: они становятся липкими, легче проникают в сосудистую стенку, хорошо захватываются макрофагами, что ускоряет развитие атеросклероза
•накоплению продуктов ПОЛ, многие из которых токсичны, канцерогенны, мутагенны (малоновый диальдегид)
•ускорению процессов старения.
На коже с возрастом увеличивается количество пигментных пятен за счет накопления пигмента – липофусцина. Он представляет собой смесь липидов и белков, связанных между собой поперечными ковалентными связями и денатурированными в результате взаимодействия с продуктами ПОЛ. Этот пигмент фагоцитируется, но не гидролизуется ферментами лизосом, и поэтому накапливается в клетках, нарушая их функции.
Процесс перекисного окисления липидов происходит и в неживых объектах. Он интенсивно протекает в пищевых продуктах, богатых ненасыщенными жирными кислотами. Именно перекисным окислением липидов объясняется порча сливочного масла, животных жиров и других продуктов питания (прогорклость).
Тесты к теме: «Биосинтез ВЖК, ТАГ и сложных липидов»
Часть 1 1. Для начала синтеза жирной кислоты необходимы субстраты:
1) |
ацил-КоА |
2) ацетил-КоА |
3) малонил-КоА |
4) НАДН+Н+ |
5) ацилкарнитин |
|
|
2.Процесс синтеза жирной кислоты:
1)носит линейный характер
2)происходит при участии карнитинацилтрансферазы I
3)носит циклический характер
4)катализирует пальмитоилсинтаза
5)активируется при физической нагрузке
3.Синтез жирных кислот увеличивается при:
1)повышении концентрации глюкозы в крови после еды
2)повышении секреции инсулина
3)увеличении секреции глюкагона
4) физической нагрузке |
5) стрессе |
|
4. Фермент АсКоА карбоксилаза активируется в присутствии: |
||
1)глюкагона 2)адреналина |
3)инсулина |
4)цитрата 5)пальмитата |
5.Координированную регуляцию синтеза и распада жирных кислот обеспечивают ферменты:
1) |
пируваткарбоксилаза |
4) карнитинацилтрансфераза I |
2) |
ацил-КоАсинтетаза |
5) цитратсинтаза |
3) ацетил-КоА-карбоксилаза
6.Соотношение скорости ферментативных реакций β- окисления и синтеза жирных кислот уровнем:
1) вазопрессина 2)кетоновых тел 3) глюкозы
4) гормонов инсулина и глюкагона 5) натрийуретического фактора
7.Непосредственными субстратами для синтеза ТАГ в печени и жировой ткани являются:
1) |
ацетил-КоА |
2) ацил-КоА |
3) глицерол-3-фосфат |
4) |
α-МАГ |
5) НАДФН |
|
8. Для синтеза ТАГ в печени могут использоваться: |
|||
1) |
холестерин |
2)глицерол |
3) ß-МАГ |
4) |
НАДФН |
5) диоксиацетонфосфат |
|
9.Метаболический путь синтеза жиров в печени и жировой ткани:
1)полностью одинаков
2)одинаков, за исключением разных путей образования α-глицерофосфата
3)приводит к синтезу частично специфичных для организма ТАГ
4)обеспечивает синтез ТАГ, полностью специфичных для организма человека
5)не зависит от влияния инсулина и глюкагона
10.Запасы триацилглицеридов в адипоцитах;
1)образуются в абсорбтивный период
2)образуются в постабсорбтивный период
3)используются как источник энергии в период голодания
4)являются компонентами клеточных мембран
5)не расходуются при голодании
11.В печени глицерол-3-фосфат может образоваться:
1)из ацетил-КоА
2)окислением дигидроксиацетонфосфата
3)фосфорилированием свободного глицерола
4)под действием фермента глицеролкиназы
5) дефосфорилированием фосфатидной кислоты
12.Из-за отсутствия в жировой ткани фермента - глицеролкиназы, глицерол-3-фосфат образуется:
1)восстановлением дигидроксиацетонфосфата
2)фосфорилирования свободного глицерола
3) |
при гидролизе ТАГ |
4) окисления 1,3-дифосфоглицерата |
5) |
при участии ферментаглицерол-3-фосфатдегидрогеназы |
|
13.Триацилглицериды, синтезированные в печени:
1)являются компонентами мембран гепатоцитов
2)упаковываются в ЛПОНП
3)секретируются в кровь
4)включаются в состав ЛПВП незрелых
5)не специфичны для организма человека
14.Начальные этапы синтеза глицерофосфолипидов и жиров:
1)имеют общие субстраты
2)протекают одинаково до образования фосфатидной кислоты
3)отличаются реакциями образования лизофосфатидной кислоты
4)протекают в митохондриях клеток
5)требуют наличия липотропных факторов
15.Активация полярного соединения используется для синтеза:
1) |
холестерина 2) |
фосфатидилэтаноламина 3)триацилглицеринов |
4) фосфатидилхолина |
5) эфиров холестерина |
|
16.Синтез фосфолипидов в организме человека может проходить через активацию:
1) |
фосфатидной кислоты |
2)глюкозы |
3) полярного соединения |
4) β-окисления |
5) перекисного окисления |
||
17.Путь синтеза фосфолипидов через активацию фосфатидной кислоты используется для образования:
1) кардиолипина |
2)фосфатидилинозитола |
3) фосфатидилэтаноламина |
4) холестерина |
5) фосфатидилхолина |
|
18.Сурфактант: |
|
|
1)внеклеточный липидный слой выстилающий поверхность лёгочных альвеол со стороны воздухоносных путей
2)в качестве основного компонента содержит дипальмитоилфосфатидилхолин
3)начинает формироваться в постэмбриональный период
4)синтезируется почками
5)содержит большое количество гликозамингликанов
19.Лептин (от лат. Leptos - тонкий, худой):
1)гормон гипоталамуса
2)в виде предшественника синезируется в печени
3)синтезируется в адипоцитах
4)имеет рецепторы в гипоталамусе и в тканях репродуктивной системы
5)повышает выработку нейропептида Y
20.Биологическое действие лептина:
1) снижение аппетита |
2)усиление липолиза 3) повышение аппетита |
4)усиление липогенеза |
5) снижение репродуктивной функции |
Часть 2 1. При недостаточном поступлении с пищей эссенциальных (незаменимых)
жирных кислот в организме нарушается синтез следующих веществ:
1) |
мочевины |
2) |
желчных кислот |
3) простагландинов |
|
4) |
глюкозы |
5) |
лейкотриенов |
6) тромбоксанов |
|
2. Ацетил-КоА является предшественником |
|
||||
1) |
глицерола |
|
2) жирных кислот |
3) холестерина |
|
4) |
аминокислот |
|
5) кетоновых тел |
6) ПВК |
|
3.Характеристика мультиферментного комплекса – синтазы ВЖК:
1)состоит из 2-х протомеров и ацилпереносящего белка (АПБ)
2)каждый из протомеров состоит из 7 доменов (ферментов)
3)расположен в матриксе митохондрий
4)катализирует синтез линоленовой кислоты
5)активируется глюкагоном
6)индуцируется инсулином
4.Наиболее активно синтез ТАГ происходит в цитозоле клеток:
1) |
миокарда |
2) почек |
3) желудка |
4) печени |
|
5) |
жировой ткани |
6) лактирующей молочной железы |
|||
5.Жировое перерождение печени предотвращают липотропные вещества, к которым относятся:
1) |
аланин |
2) метионин |
3) глюкоза |
4) |
фосфопиридоксаль |
5) кобаламин |
6) холевая кислота |
6. В регуляции депонирования и мобилизации ТАГ участвуют гормоны
1) |
инсулин |
2) вазопрессин |
3) глюкагон |
4) адреналин |
5) |
альдостерон |
6) гастрин |
|
|
7.Инсулин в жировой ткани активирует
1)синтез ЛП-липазы и ее экспонирование на поверхности капилляров
2)распад гликогена
3)транспорт глюкозы внутрь адипоцитов
4)окисление ВЖК в процессе β-окисления
5) мобилизацию ТАГ 6) гликолиз
8. Для биосинтеза некоторых фосфолипидов, в отличие от синтеза ТАГ, требуется
1)активация высших жирных кислот
2)активация фосфатидной кислоты
3)фосфорилирование азотистых оснований
4)транспорт субстратов в составе альбуминов
5)использование двух макроэргов АТФ и ЦТФ в процессе синтеза
6)этерификация только насыщенными ВЖК
9.Роль сурфактанта:
1)уменьшает поверхностное натяжение жидкости в альвеолах
2)предотвращает слипание стенок альвеол во время выдоха
3)препятствует чрезмерному растяжению альвеол при вдохе
4)приводит к развитию респираторного дистресс-синдрома
5)обладает ферментативной активностью
10.К липотропным факторам, ускоряющим синтез фосфолипидов относятся
соединения: |
|
|
1) витамин В12 |
2) витамин А |
3) холестерин |