

3.8.7. Пример: проказа в Индии
В качестве примера опишем применение близнецового метода для исследования проказы в Индии [608]. Это заболевание вызывается Mycobacterium leprae (бацилла Хансена). Известно, что не каждый, кто подвергается опасности заражения, действительно оказывается зараженным проказой, и не у всех, кто заразился, обнаруживаются клинические симптомы. Кроме того, инфекция приводит к разным последствиям в зависимости от иммунного статуса организма. У одних больных симптомы ограничиваются появлением депигментированных и безболезненных пятен (туберкулоидная проказа), а у других можно обнаружить инфильтраты (лепроматозная проказа).
Очевидные различия в восприимчивости имеют много причин; несомненно и влияние генетических факторов. Известны два типа данных, свидетельствующих об этом: накопление одной и той же формы проказы среди близких родственников и расовые различия в относительной частоте разных форм проказы. У белых и негров чаще встречается туберкулоидная форма, а на Востоке превалирует лепроматозная проказа. Близнецовые исследования туберкулеза также указывают на важность генетических факторов в восприимчивости к инфекции. Результаты других исследований, проведенных на малых выборках
3. Формальная генетика человека 285
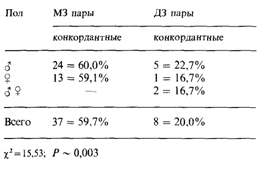
Таблица 3.29. Конкордантность 102 близнецовых пар (62 МЗ и 40 ДЗ) с проказой (Chakravartti, Vogel, 1973 [608]) |
|
больных, хотя и не полностью удовлетворительны с методической точки зрения, все же позволяют предполагать участие генетических факторов в патогенезе проказы [608].
Обсуждаемое здесь близнецовое исследование было проведено в эндемичных по проказе областях Индии - в штатах Западная Бенгалия и Андра Прадеш, где поражены по крайней мере 2-4% населения. Были предприняты определенные усилия, чтобы зарегистрировать всех близнецов, страдающих проказой в этих районах. Сначала всем, кто находился в постоянных или временных лепрозориях, задавали два вопроса: а) «являетесь ли вы близнецом?» и б) «есть ли близнецовые пары в вашей семье или деревне?». Затем исследование было распространено на деревни. Были обследованы 102 близнецовые пары по крайней мере с одним пораженным проказой.
Данные табл. 3.29 показывают, что степень конкордантности МЗ близнецов значительно выше, чем ДЗ близнецов. Кроме того, во многих пораженных МЗ парах течение болезни и размер пораженных участков обнаруживали большое
Таблица 3.30, Конкордантность и дискордантность МЗ и ДЗ близнецов по типу проказы [608]. (включены только близнецовые пары, конкордантные по проказе) |
|||
|
Конкордантность по типу |
Дискордантность по типу |
Общая сумма |
МЗ близнецы |
32 |
5 |
37 |
ДЗ близнецы |
6 |
2 |
8 |
Общая сумма |
38 |
7 |
45 |
сходство. Внутрипарные различия по возрасту начала заболевания во всех конкордантных (МЗ и ДЗ) близнецовых парах оказались меньше у МЗ, чем у ДЗ близнецов.
Поскольку проказа характеризуется разными клиническими проявлениями, то возможен анализ конкордантности и в отношении ее клинических форм. Данные табл. 3.30 показывают, что из 37 МЗ пар, конкордантных по проказе, пять оказались дискордантными по форме заболевания: один близнец имел туберкулоидную форму, а другой - лепроматозную. Именно эти пары дают благоприятную возможность для получения новых данных. Существовало мнение, что в случае лепроматозной формы проказы имеет место простой тип наследования. В качестве возможного объяснения предполагалось снижение функции Т-лимфоцитов. Однако обнаружение пяти МЗ близнецовых пар, конкордантных по проказе, но дискордантных по клинической форме заболевания, делают все эти предположения маловероятными. Таким образом, близнецовые исследования помимо выяснения того, как генетическая изменчивость популяции влияет на восприимчивость к заболеванию, позволяют проверить достаточно конкретные гипотезы о его патогенезе.
Необходимо также рассмотреть возможные смещения. Были предприняты определенные усилия, чтобы зарегистрировать в изучаемых районах всех близнецов, страдающих проказой. Однако, как показывают относительные частоты, регистрация МЗ близнецов была намного полнее, чем ДЗ близнецов. (В индийской популяции отношение МЗ/ДЗ практически такое же, как в европейских популяциях.) Как было выяснено, причина такого смещения заключалась в специфических жизненных условиях в этой части Индии. Большинство обследованных из сельских районов было неграмотно, многие не знали даже свой точный возраст. Близнецовую пару выявляли обычно только тогда, когда уже нельзя было не обратить внимание на сходство. При таких обстоятельствах ДЗ близнецов часто даже не замечали. Иногда сами сибсы не подозревали, что они близнецы.
Как неполная регистрация ДЗ близнецов могла повлиять на результат? Поскольку при регистрации предпочтение нередко отдавалось конкордантным парам, то различия между МЗ и ДЗ парами скорее могли быть недооценены. Однако более важен другой вопрос: полностью ли зарегистрированы МЗ пары? Скорее всего - нет: некоторые пары могли успешно скрыть свое заболевание, поскольку жили в колониях для нищих и находились вне поля зрения медиков; близнецы из высших социальных слоев могли
286 3. Формальная генетика человека
избежать обследования благодаря тому, что лечились у частных докторов; некоторые больные могли дать неправильные ответы, чтобы исключить социальное клеймо «прокаженных» для своих отчаявшихся близнецов или родственников. Поскольку большинство из этих факторов касается как МЗ, так и ДЗ близнецов независимо от конкордантности или дискордантности, то вряд ли селекция пробандов сильно влияла на степень конкордантности в МЗ парах. Тем не менее показатели конкордантности все же могут быть завышенными.
Что касается средовых факторов риска, то анализ дискордантности МЗ пар подтвердил, что наиболее важен непрерывный и интенсивный контакт с инфицированными. Следовательно, столь же высокие показатели конкордантности можно ожидать лишь в тех районах, где проказа высокоэндемична. Другими словами, если инфекция почти повсеместна, то вероятность развития болезни зависит главным образом от наследственной восприимчивости. В популяциях с более низкой частотой проказы инфицирование больше зависит от случайных факторов. Вот почему в этом случае можно ожидать более низкий уровень конкордантности МЗ близнецов.
Аналогичные результаты были получены при исследовании туберкулеза [919]. По данным ранних работ уровни конкордантности близнецов оказались приблизительно такими же, как и в описанном исследовании проказы. Однако эти данные были получены в то время, когда почти каждый житель индустриально развитых регионов, таких, как Европа и США, подвергался воздействию инфекции (о чем свидетельствовали положительные реакции на туберкулез). В более позднем исследовании показатели конкордантности оказались ниже [873], между тем риск заражения туберкулезом заметно снизился.
При таких соматических заболеваниях, как проказа, установленные уровни конкордантности близнецов (и, следовательно, выводы относительно степени генетической детерминации подверженности) справедливы только для тех средовых условий, в которых обитают исследуемые близнецы. При экстраполяции этих выводов на другие популяции необходимо детально рассмотреть условия жизни в этих популяциях. Например, в Центральной Европе проказа исчезла в XVII, XVIII столетиях без всякого лечения, только благодаря улучшению жизненных условий. Генетические изменения, вероятно, лишь в очень малой степени влияли на исчезновение проказы (а может быть, не влияли вовсе).