

3.6.1.4. Генетический анализ на уровне количественного фенотипа – биометрический уровень
Аддитивная модель. Во многих случаях фенотипическая изменчивость настолько сложна, что эффекты отдельных мутаций уже нельзя идентифицировать и приходится мириться с генетическими выводами (основанными на анализе сходства между родственниками), представленными в очень общем виде. Тем не менее применяемые в этих случаях «мультифакториальные» модели имеют вполне определенные характеристики, и что важнее, формулируемые на их основе прогнозы оказываются справедливыми при тестировании на реальном материале.
В простейшей из возможных моделей предполагается совместное действие нескольких генов. Предположим, что аллель, обозначаемый заглавной буквой (А или В, но не а или b), вносит свой вклад в величину признака («положительный» аллель), тогда как аллель, обозначаемый маленькой буквой (а или b, но не А или В), действует как «нулевой», т. е. не оказывает никакого эффекта на величину признака («отрицательный» аллель). Тогда фенотипическое проявление признака будет находиться в градуальной зависимости только от относительного количества положительных (или отрицательных) аллелей, вклады которых предполагаются в этой модели равными и аддитивными. Таким образом, генотипы AABBccdd..., AaBbCcDd или aabbCCDD... фенотипически не различаются (аддитивная полигения). Эту модель мы используем в дальнейшем для разъяснения ряда концепций. Необходимо пояснить, что эта модель представляет собой абстракцию и является очень упрощенной. На самом деле вклады генов, действующих в мультифакториальной системе, почти всегда будут различаться как в количественном, так и в качественном отношении. Какие-то гены окажутся более важными.
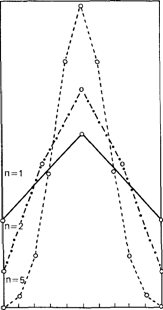
Давайте далее предположим, что рассматриваемые п диаллельных генов распространены в популяции с частотами р (для положительных аллелей) = q (для отрицательных аллелей) = 0,5. Тогда распределение фенотипических классов на произвольной количественной шкале задается биномиальной формулой (р + q)2n (рис. 3.55). Чем больше число генов , тем больше индивидов находится в центральной (т.е. ближе к средним значениям) области распределения. На первый взгляд эта зависимость может дать критерий оценки количества генов, детерминирующих признак (для чего следует сравнить эмпирическое распределение с некоторым набором теоретических). Однако такое предположение будет справедливым лишь в том случае, если каждый ген вносит в величину признака на периферии распределения точно такой же вклад, как и в центре. Это предположение можно оспорить на основе общей и часто биологически приемлемой гипотезы, согласно которой на периферии распределения дальнейшие отклонения в том же направлении достигаются труднее. Например, в случае биологически активных ве-
|
Рис. 3.55. Распределение генотипов в соответствии с биномиальным распределением (р + q)2n при р = q = 0.5 для 1, 2, 5 пар генов (п = 1,2, 5). Ось абсцисс соответствует значениям измеряемого признака. |
3. Формальная генетика человека 239
ществ и ферментов отрицательные значения активности не существуют.
Подобные рассуждения использовали для определения числа генов, детерминирующих пигментацию кожи. По нашему мнению, весьма вероятно, что число таких генов не очень велико, поскольку среди детей в браках между мулатами (т. е. множественными гетерозиготами) нередко наблюдается выщепление как чисто «белых», так и чисто «черных» индивидов.
Все распределения на рис. 3.55 имеют только одну моду (т. е. они унимодальны). Кроме того, они сходны с «нормальным» распределением. Это сходство увеличивается с возрастанием числа рассматриваемых генов (и), т. е. при возрастании п нормальное распределение является предельным случаем биномиального. Можно показать, что эта аппроксимация становится удовлетворительной как раз тогда, когда частоты положительных и отрицательных аллелей не равны. Чем ближе к симметрии, тем большие значения п требуются для достижения той же степени аппроксимации. Вообще, унимодальное распределение, которое более или менее точно аппроксимируется нормальным, является типичным для генетической модели аддитивной полигении. Однако ни унимодальность распределения, ни его форма не зависят от конкретных свойств этой модели (равных и аддитивных вкладов генов) и потому могут служить индикаторами мультифакториального наследования в более общем смысле.
С другой стороны, как показано в разд. 3.6.1.3, эти свойства не исключают наличия эффекта «главного гена» с простым типом наследования, который действует на фоне аддитивно-полигенной системы. Биологически вполне правдоподобно, что лишь несколько главных генов могут быть основными генетическими факторами ряда заболеваний, проявляющими свои эффекты на фоне многих генов, менее значимых для патогенеза этих заболеваний.
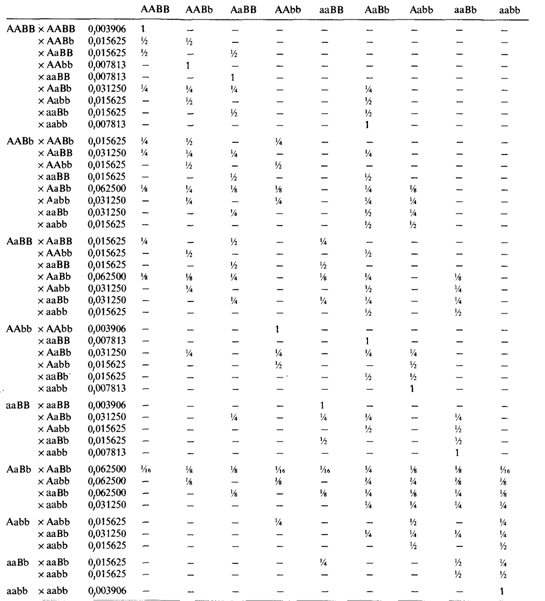
Самое первое условие установления унимодальности распределения, как и его близости к нормальному,- возможность измерить признак на какой-либо количественной шкале. Например, всех взрослых мужчин можно разбить по росту на два альтернативных класса: на тех, которые выше 167 см, и на тех, которые ниже 167 см. При такой ограниченной информации нетрудно показать на основе семейных данных, что изменчивость роста человека зависит от доминантного гена с неполной пенетрантностью. Этот пример тривиален, и аргументация очевидна, однако литература все еще полна примеров такого типа ошибок. Генетическая гипотеза не может основываться исключительно на популяционном распределении признака. Необходимы также семейные данные. Какого типа семейные данные предсказывает модель? Мы рассмотрим этот вопрос в простейшем случае двух генов с двумя аллелями каждый (А, а и В, b), действующими аддитивно и одинаково. Пусть частоты аллелей равны p1, p2 и q1 и q2 соответственно. Тогда мы будем иметь девять разных генотипов и пять разных фенотипов. Их частоты приведены в табл. 3.13. Можно вычислить частоты возможных типов браков и распределение генотипов детей для каждого типа брака родителей. Для частного случая, когда частоты всех аллелей равны 0,5 (последний столбец в табл. 3.13), вычисления приведены в табл. 3.14. Из этих распределений генотипов можно получить со-
Таблица 3.13. Генотипы и фенотипы при аддитивном полигенном наследовании |
|

240 3. Формальная генетика человека
Таблица 3.14. Типы браков, их частоты и сегрегационное отношение среди детей в случае двух генов (p1 =p2 = q1 = q2 = 0,5) |
|
3. Формальная генетика человека 241
Таблица 3.15. Распределение детей при аддитивном полигенном наследовании (не приведены пять классов 0 х —2, 0 х — 4, — 2 х — 2, — 2 х — 4, — 4 х —4, расчеты для которых могут быть проведены по тому же правилу) |
|
ответствующие фенотипические распределения детей ( табл. 3.15).
Изучаемая модель обладает следующими свойствами:
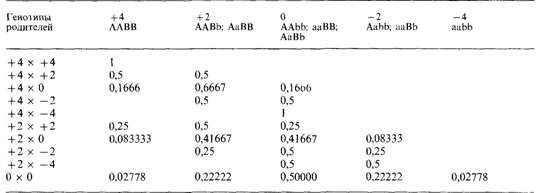
а) все получающиеся распределения имеют примерно одинаковую форму: они симметричны и унимодальны;
б) если родительские фенотипы совпадают, то средняя детей равна родительскому фенотипу. Если родительские фенотипы различны, то средняя детей точно равна среднему родительских значений (значение среднего родителя);
в) с увеличением гетерозиготности родителей ожидаемая дисперсия детей становится больше. Она наибольшая для брака 0x0 и равна 0 для браков +4x4; + 4 х —4; —4 х —4;
д) средняя детей всех лиц с одинаковым генотипом (например, дети всех лиц с фенотипом + 4) отклоняется вдвое меньше от популяционной средней, чем фенотип этих родителей (например, для родителей с фенотипом + 4 фенотипическая средняя детей равна +2).
Эта модель очень частная и простая. Но даже ее анализ оказывается уже крайне громоздким. Для изучения общего случая (п генов, частоты аллелей p1,...,pn; q1, …q2) метод необходимо видоизменить. Сначала предположим, что пара аллелей гетерозигот Аа имеет фенотипический эффект а, гомозигот АА -2α, а гомозигот aa-0. Таким образом, мы снова предположили, что гетерозиготы занимают промежуточное положение между гомозиготами. Среднее значение и дисперсию признака х можно вычислить следующим образом:
|

Наши рассуждения, которые для простоты ограничены случаем одного гена, справедливы и в общем случае п генов.
Рассмотрим теперь соотношения между родителями и детьми, а также между сибсами. Для
Таблица 3.16. Частоты пар родитель-ребенок (комбинации отец-ребенок или мать - ребенок) в популяции человека при случайном скрещивании (см. текст)
|

242 3. Формальная генетика человека
упрощения вычислений предположим, что а равна 1, тогда фенотипическое значение гомозигот АА = 2, гетерозигот Аа = 1 и гомозигот аа = 0. В табл. 3.16 показаны частоты всех возможных комбинаций мать - ребенок. Их можно объяснить следующим образом. Частота матерей АА среди всех матерей равна р2. Каждый из их детей получаст один аллель А. Вероятность, что этот аллель встретит в зиготе другой аллель А, равна р. Это дает общую частоту р2 · р = р3. Для других материнских генотипов можно провести аналогичные расчеты. Общее распределение для всей популяции (родителей и детей) будет, конечно, р2 + 2pq + q2 (маргинальные суммы в табл. 3.16).
Обозначим теперь изучаемый признак у родителей х1 , а у детей х2. Тогда полученные выше уравнения (3.5) и (3.6) дадут
Таблица 3.17. Рост родителей и взрослых детей. (По Johannsen, 1926 [726].) (Данные приведены в дюймах: 1 дюйм = 2,54 см.) |
||||||||
Рост среднего родителя |
Рост детей |
|||||||
|
|
|
|
|
|
|
|
|
|
60,7 |
62,7 |
64,7 |
66,7 |
68,7 |
70,7 |
72,7 |
74,7 |
64 |
2 |
7 |
10 |
14 |
4 |
— |
— |
— |
66 |
1 |
15 |
19 |
56 |
41 |
11 |
1 |
— |
68 |
1 |
15 |
56 |
130 |
148 |
69 |
11 |
— |
70 |
1 |
2 |
21 |
48 |
83 |
66 |
22 |
8 |
72 |
— |
— |
1 |
7 |
11 |
17 |
20 |
6 |
74 |
— |
— |
— |
— |
— |
— |
4 |
— |
|
5 |
39 |
107 |
255 |
287 |
163 |
58 |
14 |
|
Ковариацию между родителями и детьми можно получить из табл. 3.17. В общем случае ковариация двух переменных х1 и х2 определяется так: |
|

Этот важный результат был получен Фишером (1918) [664]. В случайно скрещивающейся популяции и при аддитивном действии генов коэффициент корреляции между родителем и ребенком составляет 0,5. Точно таким же способом можно показать, что коэффициент корреляции между полными сибсами тоже равен 0,5. Коэффициенты корреляции не зависят от частот аллелей р1 и q1 Такой результат означает, что родители и дети, а также сибсы имеют 50% общих генов.
Положение становится намного сложнее, если А доминирует над а. В этом случае коэффициент корреляции зависит от частот аллелей. Корреляция родитель—ребенок уже не равна корреляции сибс—сибс, а всегда меньше, за исключением случая q = 1.