

3.6.1.2. Анализ продукта гена: биохимический уровень
В этом случае идентифицировать мутантный сайт внутри гена невозможно, можно только идентифицировать ген, в котором произошла мутация. Для этого существует несколько способов.
1. Специфические белки можно охарактеризовать биохимически. Обнаруженные различия отражают генетическую изменчивость. Если белок состоит из нескольких полипептидных цепей, можно идентифицировать мутантный полипептид.
2. Многие белки выполняют функции ферментов, катализирующих специфические метаболические реакции. Следовательно, если продемонстрирован конкретный генетический блок, определен дефект фермента и исключены все другие биохимические объяснения, можно сделать вывод о мутации в конкретном гене, кодирующем данный фермент. Следующий шаг будет состоять в детальном изучении фермента.
3. Идентифицировать мутантный ген можно и на основании анализа антигенного профиля клеточной поверхности. Примерами служат группы крови и система HLA (разд. 3.5.5). В этих случаях удается выявить не только конкретные гены, но иногда также и структурные различия внутри отдельных генов.
Результаты анализа на биохимическом уровне у человека сопоставимы с результатами, полученными в экспериментальной генетике таких видов, как Drosophila melanogaster, мышь, кукуруза, шелковичный червь и другие. У этих видов во многих случаях мутации идентифицировали не на основе изменений специфических белков, ферментативных дефектов или аберрантных антигенов, а благодаря опытам по скрещиванию и рекомбинационному анализу, что в совокупности обеспечивает эффективный альтернативный подход к идентификации индивидуальных генов.
Интересно проследить, как чисто формальная концепция гена, долгое время господствовавшая в умах экспериментальных генетиков, влияла на развитие этой фундаментальной науки. Отметим, что на ранних этапах биохимическая генетика человека развивалась более успешно, нежели на других видах. Сочетание биохимического и генетического подходов к генетике бактерий и грибов привело к значительным успехам. Однако в сравнении с другими видами млекопитающих биохимическая генетика человека продолжает оставаться впереди. С появлением новых молекулярных методов генетический анализ у человека значительно упрощается.
3.6.1.3. Качественный феногенетический анализ: простые типы наследования
В данном случае выводы основаны на анализе фенотипических различий, отдаленных от первичного действия генов. Тем не менее иногда соответствие генотипа и фенотипа
232 3. Формальная генетика человека
|
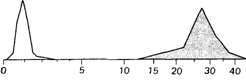
Рис. 3.46. Уровень фенилаланина в плазме крови здоровых людей и больных фенилкетонурией (окрашено), выраженный в мг%. (По Penrose, 1951.) |
столь однозначно, что напрашивается вывод о простом менделевском наследовании. Однако мы не можем с уверенностью утверждать, что этот ген единственный, поскольку один и тот же фенотип даже с одинаковым типом наследования может быть обусловлен мутациями в нескольких разных локусах.
Редко встречающиеся качественные отклонения от нормы. Эта категория охватывает большинство наследственных заболеваний. Например, индивид либо имеет нормальную пигментацию, либо утратил кожный пигмент (альбинизм). Если выражение признака можно оценить количественно, например измерить уровень метаболитов крови или мочи, то распределение фенотипических значений имеет две моды. Результаты подобного рода позволяют заподозрить наличие ферментативного дефекта и, возможно, даже идентифицировать соответствующий ген. Примерами могут служить повышенное выделение гомогентизиновой кислоты в моче больных алкаптонурией (врожденная ошибка метаболизма по Гэрроду) или повышенное содержание фенилаланина в сыворотке крови больных фенилкетонурией (рис. 3.46). Эти признаки редкие, и когда их подвергают детальному анализу, клинически сходные заболевания с одинаковым типом наследования часто оказываются генетически гетерогенными. Каковы критерии гетерогенности?
1. Если ребенок двух гомозигот с рецессивным признаком имеет нормальный фенотип, это означает, что его родители гомозиготны по разным рецессивным генам.
2. При анализе родословных в одних семьях обнаруживается тесное сцепление заболевания с геном-маркером, тогда как в других семьях эти гены сегрегируют независимо. Примером может служить сцепление одного из локусов доминантного эллиптоцитоза с геном Rh в хромосоме 1.
3. В ряде клинически сходных случаев биохимический анализ выявляет дефекты различных белков (или ферментов). Оказалось, что признаки, которые сначала считались однородными, имеют разную генетическую основу (примеры: гемофилия А и В, болезни накопления гликогена, наследственные гемолитические анемии.)
Широко распространенные признаки: бимодальное распределение. Для этой группы признаков характерно отсутствие четкого распределения фенотипов по двум классам: не каждого индивида можно с уверенностью отнести к какому-то определенному классу, поскольку возможны перекрывания. Но если область перекрывания фенотипических значений относительно небольшая, то общее распределение остается все же бимодальным.
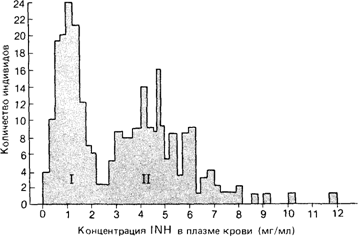
Это явление можно проиллюстрировать примером из фармакогенетики. После приема одинаковой дозы противотуберкулезного препарата - гидразида изоникотиновой кислоты (INH) его концентрация в плазме разных индивидов оказывается различной, а общее распределение имеет две моды (рис. 3.47). Можно предположить, что механизм биотрансформации лекарства детерминирован одним геном. Эта гипотеза была подтверждена в семейных исследованиях: у гомозигот Acs/Acs (Acs- аллель медленной инактивации) обнаружился высокий уровень лекарства в крови, тогда как у гетерозигот Acs/Acr (Асr – аллель быстрой инактивации) и у гомозигот Асr/Асr выявлялся низкий уровень лекарства. Был сделан вывод о том, что эти фенотипические различия в биотрансформации обусловлены наследуемыми вариантами фермента N-ацетилтрансферазы. Таким образом, подтверждение генетической гипотезы было получено благодаря открытию бимодального распределения концентраций лекарств в крови.
3. Формальная генетика человека 233
Рис. 3.47. Концентрация изониазида (INH) в плазме крови 267 членов 53 семей. Антимода полученного бимодального распределения соответствует области между 2 и 3 мг% [196]. |
|
Если исследуемый признак допускает измерение и обнаруживает бимодальное распределение, это еще не может служить доказательством простого типа наследования. Нужны более убедительные факты, поскольку возможны и другие причины бимодальности.
1. Бимодальное (или мультимодальное) распределение особенно вероятно при наличии так называемого порогового эффекта (разд. 3.6.2). Кроме того, даже в отсутствие порогового эффекта бимодальное распределение может возникать вторично, если признак обнаруживает тенденцию к «самоусилению» при достижении определенной степени выражения. Одним из примеров может служить кровяное давление: при определенном повышении его уровня почки могут уже не справляться со своими функциями, что влечет за собой более резкое повышение кровяного давления. Бимодальность может имитироваться также особенностями регистрации семейного материала.
2. Средние значения двух распределений могут быть столь близкими друг к другу, что бимодальность окажется незаметной. Харрис и Смит [703] исследовали условия, при которых комбинация двух нормальных распределений может привести к бимодальному распределению:
а) два нормальных распределения с одинаковыми дисперсиями дают в совокупности бимодальное распределение, только если разность средних значений по крайней мере в два раза больше общего стандартного отклонения;
б) в случае различных дисперсий, если разность между средними значениями выражать в единицах меньшего стандартного отклонения, то коэффициент должен изменяться от 2 (когда дисперсии равны) до 2,6 (когда дисперсии сильно различаются);
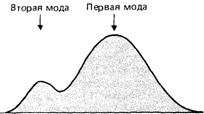
в) если средние значения настолько близки между собой, что бимодальность общего распределения не выявляется, а численности индивидов в двух распределениях различаются немного, то на наличие двух разных распределений может указывать так называемая «битангенциальность» общего распределения (рис. 3.48).
На практике трудно убедиться в наличии именно такого распределения. Встречающиеся в природе переменные редко имеют идеальное нормальное распределение, и поэтому следует помнить о случайных выборочных отклонениях. Убедиться в том, что случайные отклонения иногда искажают распределение эмпирически наблюдаемых значений при умеренном объеме выборки, можно при сравнении рис. 3.48 с рис. 3.47.
Вывод. Унимодальное распределение может свидетельствовать о моногенном наследовании, однако в отсутствие дополнительных данных отличить моногенную модель от мультифакториальной невозможно.
234 3. Формальная генетика человека
На рис. 3.49 представлен противоположный пример: распределения ферментативной активности разных электрофоретически идентифицируемых генетических вариантов GPT (глутамат-пируват-трансаминазы) четко различаются. При этом общее распределение в популяции является почти нормальным, что принято интерпретировать как отражение мультифакториальной природы признака. Тем не менее в данном случае общее распределение в популяции детерминировано лишь двумя аллелями (GPT1, GPT2) и соответственно тремя фенотипами (GPT1, GPT2-1, GPT2). Много случаев такого рода было обнаружено при анализе изменчивости ферментативной активности в различных системах генетически полиморфных ферментов (разд. 6.1).
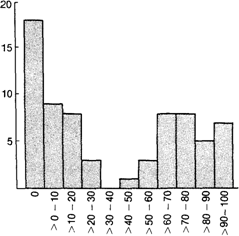
Распознавание бимодальности может быть существенно затруднено, если два класса имеют очень разные частоты, например если численность индивидов одного класса составляет менее 10% от численности другого (рис. 3.50). В этой ситуации возникает сомнение, является ли меньшая мода истинной, отражающей наличие генетически особой группы. И в таких случаях также возможны:
|
|
Рис. 3.48. Битангенциальное распределение. (Harris, Smith, 1951 [703].) |
Рис. 3.49. Распределения ферментативной активности для трех генотипов GPT и их суммарное распределение, очень сходное со скошенным нормальным [8]. |


а) случайные отклонения;
б) пороговый эффект: к этой возможности следует отнестись серьезно, особенно если фенотипические значения, образующие вторую моду, близки к нулевым.
Влияние случайных отклонений можно избежать, если исследовать большее количество индивидов, но при этом не исключается, конечно, пороговый эффект. Необходимы семейные исследования, особенно в случаях аутосомно-рецессивного или сцепленного с полом наследования. В таких семьях следует ожидать наличия двух генотипов примерно с равными частотами, и поэтому бимодальность будет выражена более четко, чем в популяционной выборке. При исследовании семей пробандов, фенотипы которых соответствуют меньшей моде, в пользу простого типа наследования свидетельствуют следующие факты (если они типичны для большинства семей):
а) четко выраженное бимодальное распределение признака в сибствах, причем значения фенотипических признаков у сибсов, соответствующие моде «нормы», имеют распределение такое же, как и в общей популяции;
б) значения у родителей также образуют бимодальное распределение с тем дополнительным условием, что по крайней мере один из родителей должен попасть в группу второй моды, отличной от «нормы»;
3. Формальная генетика человека 235
|
Рис. 3.50. Бимодальное распределение количественного признака в популяции человека. Один из двух типов распространен намного чаще, чем другой. |
в) в этих сибствах соотношение пораженных и непораженных должно быть примерно 1:1. Если оба родителя попадают во вторую моду, т. е. оба гетерозиготны, то соотношение пораженных и непораженных сибсов должно быть примерно 3:1.
При мультифакториальном наследовании ложная бимодальность может оказаться следствием порога в области нулевых значений. В результате распределение тех сибсов, фенотипические значения признака которых, можно отнести к первой моде, будет обнаруживать меньшее среднее значение, чем в популяции. Кроме того, значения признака у родителей чаще будут относиться к первой моде с более низким средним значением, чем в популяции (рис. 3.51).
Признак, для которого вывод об аутосомнодоминантном наследовании был сделан на основе указанных критериев, - это низкая амплитуда электроэнцефалограммы (ЭЭГ) [921]. Человеческий мозг постоянно порождает электрические колебания, которые после прохождения через соответствующие усилители можно записать в виде кривых на бумаге. Для этого к голове в определенных точках прикладывается несколько (8-16) электродов. Испытуемый расслабляется и отдыхает, но не спит (типы ЭЭГ сна различны и сами по себе часто используются в специальных диагностических целях). В ЭЭГ отдыхающего здорового взрослого человека обнаруживается несколько разных типов волн, среди которых наиболее примечательны а-волны, характеризующиеся умеренной частотой колебаний. Кроме этого могут присутствовать быстрые β-волны (с частотой более 13 колебаний в секунду) и несколько видов медленных θ-волн (4-8 колебаний в с) (рис. 3.52). Комбинации различных элементов кривой ЭЭГ многообразны, их можно сравнивать между собой. Почти каждый человек имеет свою собственную, характерную тольку для него ЭЭГ, которая остается постоянной многие годы в отсутствие таких заболева-
|
|
Рис. 3.51. Бимодальное распределение в случае, когда вторая мода близка к нулю. Различие между простым диаллельным наследованием и мультифакториальной моделью. Отметим, что при мультифакториальном наследовании первая мода сдвинута влево, тогда как при диаллельном наследовании первая мода совпадает с популяционной модой и модой в тех семьях, в которых происходит расщепление на два типа. |
Рис. 3.52. Типы колебаний (волн) электроэнцефалограммы человека. |


236 3. Формальная генетика человека
ний, как эпилепсия или опухоль мозга, а также исключая такие кратковременные физиологические состояния, как сильное утомление или тяжелая интоксикация. В детстве и юности ЭЭГ развивается из нерегулярных форм с относительно медленными волнами, завершает свое развитие примерно к 19 годам, а затем меняется очень мало. Индивидуальные различия в скорости развития огромны, что объясняет высокую изменчивость ЭЭГ в детском возрасте. Близнецовые исследования показали, что в норме индивидуальные различия ЭЭГ детерминированы почти исключительно генетически.
Примерно у 4% взрослых индивидов из общей популяции обнаружены ЭЭГ со следующими характеристиками:
а) а-волны затылочной области полностью отсутствуют или выявляются только короткое время с очень низкой амплитудой;
б) ЭЭГ может выглядеть абсолютно плоской или обнаруживать нерегулярную картину с βили θ-волнами низкой амплитуды;
в) в противоположность ЭЭГ с нормальным α-ритмом отсутствует реакция на открывание глаз. После закрывания глаз могут появиться, но не обязательно, отдельные а-волны.
Первым требованием генетического анализа является, конечно, возможность количественно оценить α-ритм в затылочной области. Одной из мер может служить так называемый а-индекс, определяемый следующим образом:
![]()
На рис. 3.53 показано распределение этого индекса в 30 сибствах, зарегистрированных по пробандам с низкой амплитудой ЭЭГ. Это распределение имеет два максимума, один около 70-80, а другой около 0. Второй максимум соответствует низкой амплитуде. На первый взгляд это распределение свидетельствует в пользу моногенной гипотезы, однако второй максимум близок к нулевому значению и, следовательно, бимодальность распределения может оказаться следствием порога, расположенного в нулевой точке, т. е. фиктивной.
Имеются ли дополнительные аргументы в пользу моногенного наследования? Распределение среди родителей является бимодальным, а распределение вокруг первой моды хорошо согласуется с распределением в общей популяции. Еще важнее то, что во всех семьях, зарегистрированных по пробанду, по крайней мере один родитель имел низкую амплитуду ЭЭГ (рис. 3.54). Сегрегационный анализ данных по всем семьям с двумя пораженными родителями дал оценку вероятности расщепления, близкую к 75%, но по всем семьям с одним пораженным родителем оценка этой вероятности была значительно ниже ожидаемого значения, равного 50%. Оказалось, что такой результат - следствие включения в выборку подростков в возрасте от 10 до 19 лет: низкая амплитуда ЭЭГ как раз только формируется в течение этого периода. Ограничение анализа только лицами 19 лет и старше приводит к превосходному соответствию ожидаемых и наблюдаемых сегрегационных отношений (табл. 3.12). (Ожидаемые значения иногда выше, чем 0,5 или 0,75 соответственно, поскольку вследствие высокой частоты признака среди родителей ожидается некоторая доля гомозигот.) В описанном случае анализ распределений количественного признака (α-индекс) в семьях позволил сделать вывод об аутосомно-доминантном типе наследования.
В принципе сходные критерии можно использовать при изучении Х-сцепленного рецессивного наследования. Однако в этом случае анализ распределений может оказаться труднее для женской части популяции, поскольку ожидается тримодальное распределение: две гомозиготы и одна гетерозигота. Примером может служить активность глюкозо-6-фосфатдегидрогеназы (G6PD): так как существуют большие перекрывания между «нормой» и гетерозиготами, а также между гетерозиготами и гомозиготами по недостаточности G6PD, в женской части популяции трудно идентифицировать тримодальность.
|
Рис. 3.53. Распределение а-индекса в 30 сибствах из семей пробандов с низкоамплитудными ЭЭГ (см. текст). |
3. Формальная генетика человека 237
Таблица 2.12. Низкоамплитудная ЭЭГ ( + ): сегрегационный анализ 60 семей со 117 детьми |
|

|
Рис. 3.54. Семейное обследование. У матери и первой дочери низкоамплитудные ЭЭГ, у отца и второй дочери стандартная ЭЭГ с α-ритмом.Униполярные отведения. |

238 3. Формальная генетика человека