

2.3.6. Новая генетика и концепция гена
Молекулярная цumoгенетика. Методы новой генетики важны не только для изучения структуры генов, они оказывают все возрастающее влияние на эффективность цитогенетической диагностики. Например, мелкие хромосомные аберрации, такие, как делеции и транслокации, идентифицируются теперь намного точнее, чем в недавнем прошлом. Мелкие делеции, не выявляемые классическими методами, в настоящее время можно обнаружить, если имеется ДНКзонд для делетированного участка. Возможна также очень ранняя (8-10 недель беременности) пренатальная диагностика пола эмбриона с использованием Y-специфического ДНК-зонда, биопсии ворсин хориона (разд. 9.1) и методики Саузерна. Получены новые, впечатляющие данные относительно структуры, филогенетических взаимосвязей и эволюции обеих половых (X и Y) хромосом (разд. 3.4). Новая технология несомненно должна войти в арсенал средств тех учреждений, которые занимаются генетическим консультированием и пренатальной диагностикой. Современная служба медико-генетической помощи остро нуждается в специальном дорогостоящем оборудовании и хорошо обученном персонале. Готово ли общество к крупномасштабным затратам на такую службу? Как поступать слаборазвитым странам, у которых и без того имеется множество неотложных экономических трудностей? Возникают и совершенно новые проблемы. Например, в некоторых популяциях еще существуют традиции, согласно которым сыновей предпочитают дочерям. Не приведут ли появившиеся возможности осуществлять выбор пола детей к сложным и непредвиденным последствиям?
Что такое ген? (рис. 2.100). В классической генетике ген рассматривался как единица мутации, рекомбинации и функции. Принято было считать также, что гены расположены в хромосоме в линейном порядке подобно бусинкам на нити. Однако детальный генетический анализ показал, что такое представление является упрощенным: например, у дрозофилы два тесно сцепленных мутантных гена, будучи на одной хромосоме (т. е. в цис-положении), дают меньший фенотипический эффект, чем те же две мутации, расположенные на гомологичных хромосомах (т.е. в транс-положении). Нередко фенотипический эффект и вовсе отсутствовал. Гены, демонстрирующие такой цис-транс-эффект, были названы псевдоаллелями (см. также разд. 3.5.1; рис. 3.30). В дальнейшем биохимический анализ показал, что цис-транс-эффект возникает в том случае, когда две мутации затрагивают разные сайты в пределах структурного гена, кодирующего один простой белок. Когда две такие мутации находятся в цис-положении, гомологичный нормальный ген способен контролировать синтез функционально интактного белка. С другой стороны, когда две мутации находятся в транс-положении, интактный белок не образуется. Развитие молекулярной генетики в 50-е гг. сделало необходимым введение новой терминологии. По предложению Бензера (1957; [569]) единицу рекомбинации стали называть реконом, единицу мутации-мутоном, а единицу функции-цистроном (по цис-транс-эффекту). В последующие годы было показано, что рекон и мутон соответствуют отдельному нуклеотиду-самой маленькой единице генетического материала, тогда как цистрон соответствует фрагменту ДНК, кодирующему одну полипептидную цепь. Из этих трех терминов только последний стал популярным среди гене-
2. Хромосомы человека 149
|
Рис. 2.100. Историческое развитие концепции гена. Гены были постулированы Иогансеном. Они заменили в наших представлениях наследственные «задатки» Менделя, которые, по его мнению, ответственны за передачу признаков. Обнаружение сцепления генов в хромосомах привело к модели «бусинки на нити», согласно которой гены (бусинки) нанизаны на хромосому (нить). Материальная основа гена оставалась неизвестной. Важнейшим этапом в развитии концепции гена было открытие того факта, что за передачу наследственной информации в клетке ответственна ДНК. Геном стали считать специфическую последовательность ДНК, которая кодирует полипептидную последовательность (три нуклеотида детерминируют одну аминокислоту). Вскоре было показано, что единица, определяющая свойства полипептида (цистрон), отличается от единицы рекомбинации (рекон) и от единицы мутации (мутон). Мутон соответствует одному основанию. Затем было обнаружено, что определенные участки ДНК не кодируют белки, а выполняют, по-видимому, регуляторную роль. Было показано также, что структурные гены прерываются некодирующими последовательностями (нитронами). Кодирующие последовательности структурных генов назвали экзонами. Границы фланкирующих и «регуляторных» последовательностей (5') и (3') не уточнены. Следовательно, в середине 80-х гг. в структуре гена еще остается много неясного. Кодирующие и вставочные последовательности гена можно определить то'чно (см. также [247а]). |
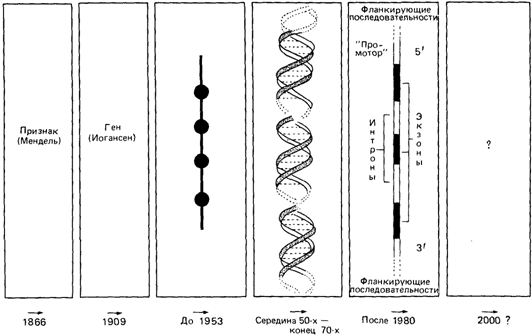
тиков как синоним гена в смысле единицы функции.
По мере углубления наших знаний о структуре генетического материала (наличие интронов, промоторных последовательностей, псевдогенов и т.д.), и особенно в связи с открытием мультигенных семейств генов, таких, как семейство Hbβ, понятие «ген» становится все более и более расплывчатым. В настоящее время все еще неясно, какие из длинных последовательностей ДНК, включая повторы типа Alu, рассеянных среди кодирующих районов, необходимы для функции гена. Хотя о локусе Hbβ мы знаем больше, чем о любом другом гене млекопитающих (см. также разд. 4.3), трудно четко определить границы этого генного комплекса. По-видимому, для определения гена разумно пользоваться цис-транс-тестом (так называемый компле-
150 2. Хромосомы человека
ментационный критерий). В анализе генетической гетерогенности на биохимическом уровне этот критерий действительно используется, хотя получаемая с его помощью информация является все же недостаточно специфичной. Для нового определения «гена» необходима дополнительная информация, основанная на результатах молекулярного анализа на уровне ДНК.
Новые результаты по структуре гена и формальная генетика. Обсуждение данных о структуре гена (разд. 2.3) может создать впечатление, что большинство результатов классического генетического анализа устарело. Это, однако, не так. Для анализа типа наследования в семьях, для решения вопроса о генетическом сцеплении между неаллельными генами, для изучения генетической структуры популяции принципы формальной генетики не только применимы, но и необходимы. Ситуацию можно сравнить с той, какая сложилась в свое время в физике: квантовая механика помогла нам понять природу света намного полнее, чем классическая физика. Однако геометрическая оптика останется не только правильной, но и совершенно необходимой для многих практических приложений, таких, как конструирование очков или микроскопов. Следовательно, она является необходимой частью каждого руководства по физике.