

2. Трансмембранная асимметрия липидов
Каждая мембрана клетки замкнута, т.е. имеет внутреннюю и внешнюю поверхности, различающиеся по липидному и белковому составам - эту особенность мембран называют трансмембранной (поперечной) асимметрией.
Липидная асимметрия возникает прежде всего потому, что липиды с более объёмными полярными "головками" стремятся находиться в наружном монослое, так как там площадь поверхности, приходящаяся на полярную "головку", больше. Фосфатадилхолины и сфингомиелины локализованы преимущественно в наружном монослое, а фосфатидилэтаноламины и фосфатидилсерины в основном во внутреннем.
Липиды в некоторых биологических мембранах с довольно большой частотой мигрируют с одной стороны мембраны на другую, т.е. совершают "флип-флоп" (от англ, flip-flop) перескоки (рис. 5-7). Перемещение липидных молекул затрудняют полярные "головки", поэтому липиды, находящиеся на внутренней стороне мембраны, имеют относительно высокую скорость

Рис. 5-6. Положение молекулы холестерола в мембране. Молекула холестерола располагается в липидном слое мембраны параллельно алифатическим цепям молекул фосфо- и гликолипидов. Гидроксильная группа холестерола контактирует с гидрофильными "головками" этих липидов.
3. Жидкостностъ мембран
Для мембран характерна жидкостность (текучесть), способность липидов и белков к латеральной диффузии. Скорость перемещения молекул зависит от микровязкости мембран, которая, в свою очередь, определяется относительным содержанием насыщенных и ненасыщенных жирных кислот в составе липидов. Микровязкость меньше, если в составе липидов преобладают ненасыщенные жирные кислоты, и больше при высоком содержании насыщенных жирных кислот.
Ацильные (алифатические) остатки ненасыщенных жирных кислот имеют так называемые "изломы" (см. раздел 8). Эти "изломы" препятствуют слишком плотной упаковке молекул в мембране и делают её более рыхлой, а следовательно и более "текучей". На текучесть мембран также влияют размеры углеводородных "хвостов" липидов, с увеличением длины которых мембрана становится более "текучей".
4. Функции мембранных липидов
Фосфо- и гликолипиды мембран, помимо участия в формировании липидного бислоя, выполняют ряд других важных функций.
Липиды формируют среду для функционирования Мембранных белков, принимающих в ней нативную конформацию. Выделенные из мембран ферменты, лишённые липидного окружения, как правило, не проявляют каталитической активности.
Некоторые мембранные липиды - предшественники вторичных посредников при передаче гормонального сигнала. Так, фосфати-дилинозитол-4,5-бисфосфат (ФИФ2) под действием фермента фосфолипазы С гидролизуется до диацилглицерола (ДАТ), активатора протеинкиназы С и инозитол-1,4,5-трифосфата (ИФ3) - регулятора кальциевого обмена в клетке (рис. 5-8). ДАГ, ИФ3, протеинкиназа С и Са2+ - участники инозитолфосфатной системы передачи сигнала.
Кроме того, некоторые липиды выполняют "якорную" функцию, например к фосфатидил-инозитолам через олигосахарид могут присоединяться специфические белки наружной поверхности

Рис. 5-8. Гидролиз фосфатидилинозитол-4,5-бисфосфата.
клетки (рис. 5-9). Фосфатидилинозитол с присоединённым к нему олигосахаридом (гликаном) называют фосфатидилинозитолгликаном. Связь белков с этой молекулой (гликаном) осуществляется через фосфоэтаноламин. Пример такого "заякоренного" белка - ацетилхолинэстераза, катализирующая гидролиз ацетилхолина в синаптической щели. Этот фермент фиксируется на поСтеинаптической мембране, ковалентно присоединяясь к фосфатидилино-зитолгликану. Под действием фосфолипазы С может происходить отделение белков от внешней поверхности клетки.
Липиды могут быть аллостерическими активаторами мембранных ферментов. Например, р-гидроксибутиратдегидрогеназа, участвующая в окислении кетоновых тел (см. раздел 8), локализована на внутренней мембране митохондрий. Каталитическая активность фермента проявляется только в присутствии фосфатидилхолина.
Фермент протеинкиназа С катализирует реакции фосфорилирования белков по аминокислотным
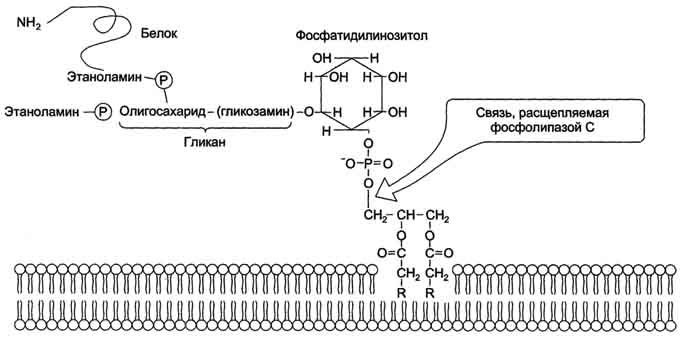
Рис. 5-9. "Якорная" функция фосфатидилинозитолгликанов.
остаткам серина и треонина. В неактивной форме протеинкиназа С находится в цитозоле. Однако после стимуляции клетки (повышение в клетке концентрации кальция) фермент быстро активируется ионами кальция и оказывается связанным с мембраной. Функционально активная протеинкиназа С - комплекс, содержащий мономер фермента, молекулу диацилглицерола, один или более ионов Са2+ и четыре молекулы фосфатидилсерина.
Креатинкиназа, фермент катализирующий образование макроэргического соединения креа-тинфосфата Для проявления его активности требуется специфическое взаимодействие с кардиолипином внутренней мембраны митохондрий.