

21.3.4. Картирование внутрикоркового взаимодействия при шизофрении
Шизофрения — заболевание, которое связано, в основном, не с поражением определенных структур мозга, а с нарушением взаимоотношений между этими структурами, с их функциональной дезинтеграцией. Электрофизиологически нарушения взаимоотношений между мозговыми структурами изучаются с помощью методов синхронизации биопотенциалов [Ливанов, 1972], корреляционного анализа [Монахов, Стрелец, 1970], когерентности [Болдырева, 1992; Weiss, Rapplesberger, 2000] и картирования внутрикорковых связей [Иваницкий, 1990].
Когерентность — степень синхронизации, основанная на оценке интеграции между различными отделами мозга на частоте, усредненной для всего частотного диапазона у каждого испытуемого. М. Н. Ливанов [1972] считал, что синхронизация создает условия для возникновения функциональной связи и, возможно, является выражением этой связи.
А
При выполнении
когнитивного задания
количество
межполушарных
связей у здоровых
увеличивается
до восьми. У больных
шизофренией обеих
групп с помощью
этого метода
удалось выявить
полное отсутствие
межполушарных
связей
Нами разработана модификация этого метода [Стрелец и др., 2000], которая заключается в том, что после нахождения пиков, совпадающих по частоте в отведениях от различных областей, связи между этими пиками не усредняются, а по вероятности их появления выделяются наиболее типичные из них; достоверность этих связей определяется по методу Монте-Карло.
21.3.5. Взаимосвязи между корковыми областями на высокочастотном бета-ритме
Наибольший интерес представляет собой проблема взаимосвязей на высокочастотном бета-ритме, поскольку на этой частоте, согласно последним данным, осуществляется взаимодействие (binding) между различными отделами мозга во время сенсорно-перцептивных процессов, при когнитивной деятельности и мышлении [Basar, 1992; Whittington et al., 1997; Traub et al, 1996] (гл. 8 ).
При шизофрении архитектура внутрикорковых связей на высокочастотном бета-ритме значительно отличаются от нормы. На рис. 21.2 приведены результаты нашего исследования когерентности у групп здоровых (верхний ряд), больных шизофренией с позитивными симптомами (средний ряд) и больных шизофренией с негативными симптомами (нижний ряд).

Рис. 21.2. Взаимосвязи на высокочастотном бета-ритме у здоровых (1), больных с «позитивными» (2) и «негативными» (3) симптомами, полученные методом когерентности в покое (А) и во время выполнения когнитивного задания (Б). Шкала в нижней части рисунка соответствует частотам, на которых обнаруживаются связи
Как видно из рисунка, у здоровых в состоянии покоя имеется только две межполушарных связи — между центральными и затылочными областями. При выполнении когнитивного задания количество межполушарных связей у здоровых увеличиливается до восьми, причем наряду с прямыми связями — между всеми гомологичными отведениями — наблюдаются также «косые» связи — между лобными отведениями каждого из полушарий с центральными отведениями противоположных полушарий, а также между левым центральным и правым теменным и между правым теменным и левым затылочным отведениями.
У больных шизофренией обеих групп с помощью этого метода удалось выявить полное отсутствие межполушарных связей на высокочастотном бета-ритме как в состоянии покоя, так и при выполнении когнитивной пробы.
Таким образом, феномен «связывания» различных, пространственно удаленных мозговых областей коры в процессе когнитивной деятельности при шизофрении нарушен. Подобные нарушения интеграции у больных шизофренией описаны в современной литературе [Bennet, 1997; Gruzelier, 1999 ], однако в этих и приведенных выше работах давались пробы на одно из полушарий, тогда как в нашем исследовании предъявлялась проба, специально направленная на исследование интеграции между двумя полушариями. Можно предполагать, что дезинтеграция, «разрыв» большого числа нейронных связей, в большей степени межполушарных, по-видимому, обусловливает неспособность больных шизофренией к адекватной активации корковых областей в условиях когнитивной деятельности.
Методом исследования внутрикорковых связей [Иваницкий, 1990] (с вычислением средней по группе частоты) были получены данные, сходные с теми, которые получены методом когерентности, однако у обеих групп больных было выявлено по одной межполушарной связи при выполнении когнитивной деятельности — между затылочными областями (рис. 21.3).
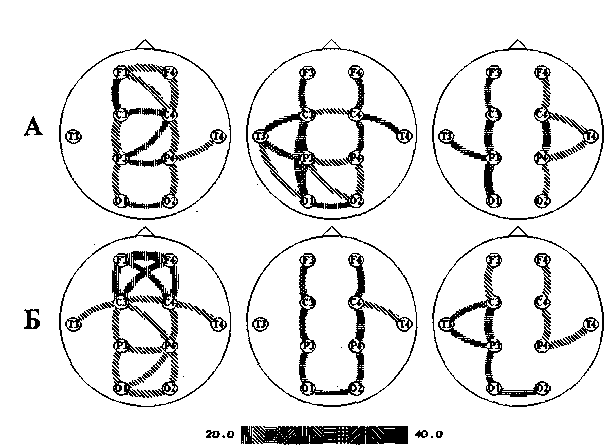
Рис. 21.3. Связи на высокочастотном бета-ритме у здоровых (1), больных с «позитивными» (2) и «негативными» (3) симптомами, полученные при картировании усредненных связей в покое (Л) и во время выполнения когнитивного задания (Б). Шкала в нижней части рисунка соответствует частотам, на которых обнаруживаются связи
И
Самые интересные
данные о характере
взаимосвязей на
высокочастотном
бета-ритме могут
быть получены
с помощью метода
определения
наиболее типичных
для каждой группы
связей; в норме
наиболее типичными
оказались связи
на частоте
приблизительно 40 ГЦ
У
больных
с
позитивной
симптоматикой
межполушарные
связи
отсутствуют
С
У больных
шизофренией
с негативной
симптоматикой
на частоте
около 40 Гц имеется
только три связи,
возникающие
в правом полушарии
Самые интересные данные о характере взаимосвязей на высокочастотном бета-ритме были получены с помощью третьего метода, определения наиболее типичных для каждой группы связей, который позволяет определить не только наличие или отсутствие связей, но и частоту, с которой они появляются, поскольку, как уже отмечалось, частота в этом методе не усредняется по группе. Этим методом было показано (рис. 21.4), что в норме наиболее типичными оказались связи на частоте приблизительно 40 Гц, которые характерны для феномена взаимодействия. В состоянии покоя на этой частоте отмечались связи между теменной и височной областями левого полушария и между центральной и затылочной областями правого; внутриполушарные связи регистрировались между левой лобной и правой центральной, а также между затылочными областями. При выполнении когнитивной пробы появлялось четыре межполушарных связи на частоте 38 Гц: правой центральной области с лобной, височной, теменной и затылочной областями и шесть внутриполушарных (в левом полушарии).
У больных с позитивной симптоматикой межполушарные связи отсутствуют. При когнитивной деятельности у них наблюдаются внутриполушарные связи на частоте около 40 Гц —- между лобной и височной областями в обоих полушариях; в правом полушарии имеются также связи височной области с центральной и теменной, теменной с лобной и центральной областями.
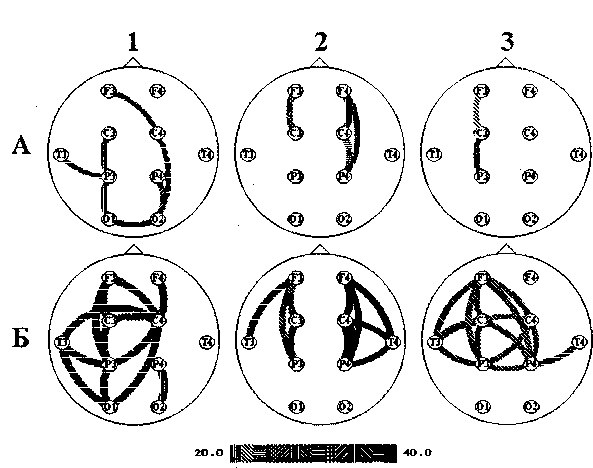
Рис. 21.4. Связи на высокочастотном бета-ритме у здоровых (1), больных с «позитивными» (2) и «негативными» (3) симптомами, полученные при картировании типичных связей в покое (А) и во время выполнения когнитивной пробы (Б). Шкала в нижней части рисунка соответствует частотам, на которых обнаруживаются связи
У больных шизофренией с негативной симптоматикой на частоте около 40 Гц имеется только три связи, возникающие в правом полушарии при когнитивной деятельности — между правой височной и лобной, центральной и теменной областями.
Таким образом, третий метод исследования внутрикоркового взаимодействия, метод исследования типичных для группы связей позволяет выявить нарушения межполушарного взаимодействия на частоте около 40 Гц у обеих групп больных шизофренией. Однако, если у острых больных с позитивными симптомами межполушарные связи вообще отсутствуют, то у больных с негативными симптомами возникает новая сеть взаимосвязей на относительно низкой частоте бета-ритма — около 30 Гц. У острых больных полное разрушение сети межполушарных связей обусловлено высокой вариабельностью физиологических показателей, которая, в свою очередь, определяет разнообразие клинических симптомов. Образование новой сети взаимосвязей на отличной от нормы частоте у больных с негативными симптомами может указывать на переход заболевания в хроническую стадию. Можно предположить, что нарушение соответствия частотных спектров двух полушарий при шизофрении, обусловленное разными уровнями их активации, препятствует межполушарной передаче и «приему» информации.
Современными исследованиями [Traub et al., 1996; Whittington et al., 1997] показано, что бета-ритм может возникать из гамма-ритма (частотой выше 40 Гц), который и обусловливает наблюдающиеся при шизофрении нарушения взаимосвязей. Согласно этим данным, гамма-ритм образуется при тетанической стимуляции гиппокампальных срезов за счет разрядов тормозных интернейронов, которые взаимодействуют друг с другом, поддерживая ритмическую импульсацию. Обнаружено, что при тетанической стимуляции, вдвое превышающей ту, которая необходима для возникновения гамма-ритма, этот ритм превращается в бета-ритм, функциональная значимость которого достаточно ясна.
Тот факт, что в норме межполушарные связи по высокочастотному бета-ритму, выполняющему активационную функцию, повышаются при когнитивной деятельности, а при шизофрении — отсутствуют или возникают на более низкой частоте, может быть связан с нарушением при шизофрении шипикового аппарата тормозных интернейронов, участвующих в генерации гамма-ритма [Whittington et al., 1997]. Несомненно, описанные выше предположительные механизмы нарушения внутрикоркового взаимодействия при шизофрении будут постоянно дополняться как электрофизиологическими, так и биохимическими данными. Так, уже известно, что образование гамма-ритма опосредуется рядом медиаторов, в частности га-мааминомасляной кислотой (ГАМК).