

20.7. Мозг, психика и эволюция генома: на пути к теории нейроэволюции
Сказанного в настоящей главе достаточно, чтобы увидеть, что мы находимся только на самых начальных подступах к решению проблемы нейроэволюции — проблемы, которая связывает системы мозга, опосредующие участие психики и сознания в процессах естественного отбора, с эмбриональным развитием нервной системы, генами и морфологической эволюцией.
Однако уже сегодня мы можем определить основные направления решения этой проблемы и те элементы, из которых должна складываться полноценная теория нейроэволюции.
Ясно, что в эволюции мозга, как и других органов тела, критическую роль играли регуляторные гены, определяющие процессы эмбрионального развития. По-видимому, к таким генам относятся прежде всего гены различных транскрипционных факторов и морфорегуляторных молекул.
Но, в отличие от других соматических органов, в созревшем мозге многие из этих генов вновь активируются — в ситуациях новизны и научения. Вследствие этой реактивации нейроны фиксируют свое участие во вновь образующихся функциональных системах за счет долговременного изменения своих синантических связей. В результате в нервной системе морфогенез по сути никогда не прекращается.
Таким образом, в отношении мозга две фазы эволюционного цикла — созревание (первичный системогенез) и адаптивные модификации (вторичный системогенез) функциональных систем, обеспечивающих дифференциальное выживание, — оказываются тесно связанными на уровне механизмов регуляции экспрессии генов (рис. 20.6).
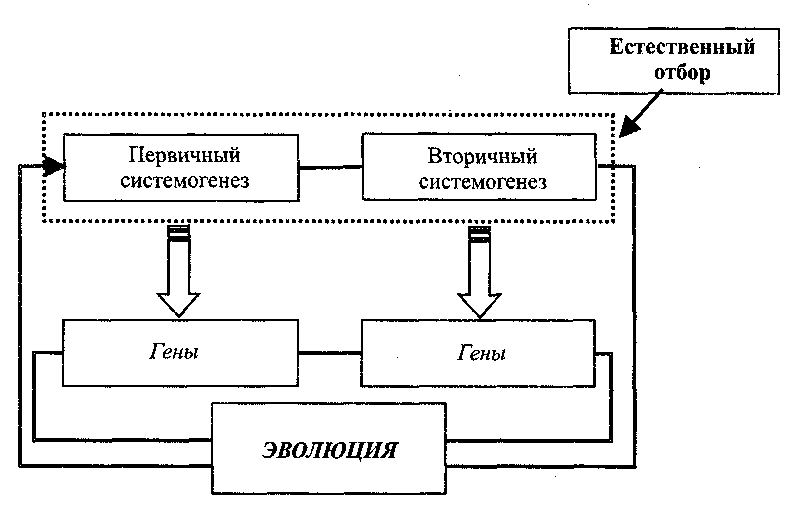
Рис. 20.6. Связь и генетическое родство процессов первичного системогенеза (созревания функциональных систем) и вторичного системогенеза (адаптивной модификации существующих функциональных систем и возникновения новых систем при научении (см. гл. 14, разд. 7.9) процессах системной специализации) в эволюционном цикле
Такое сходство естественно заставляет думать об интенсивных эволюционных взаимодействиях и переходах между двумя этими доменами.
Есть основания полагать, что именно на путях исследования этих взаимодействий может быть получен ответ на один из наиболее сложных и захватывающих вопросов современной науки — как в ходе филогенеза мозг стал органом, определяющим эволюцию генома?
В решении этого вопроса критическую роль играет то, что на стадии вторичного системогенеза экспрессия генов в мозге оказывается под контролем системных когнитивных процессов, эффективность которых постоянно оценивается естественным отбором.
Глава 21 Клиническая психофизиология
21.1. Предмет и задачи
Освещение проблем психофизиологии не может считаться достаточно полным, если оно не включает раздела, посвященного психической патологии. Еще И. П. Павлов считал, что психиатрия является как бы физиологическим экспериментом, поставленным самой природой.
С одной стороны, достижение конечной цели психиатрии, то есть понимание природы психических заболеваний, нахождение методов их диагностики и лечения, невозможно только через наблюдения, анализ и обобщение клинического материала. Необходимое понимание тех глубинных мозговых процессов, которые проявляются в психопатологии, тесно связано с развитием современных инструментальных методов исследования мозга, среди которых особая роль принадлежит картированию электроэнцефалограммы (ЭЭГ) и другим методам «нейроимиджинга», или «визуализации живого мозга». Эти методы вкратце описаны в главе 2.
С другой стороны, изучая патофизиологию когнитивных и эмоциональных процессов и сопоставляя ее с нарушениями этих функций при шизофрении и депрессии, можно обнаружить некоторые нейрофизиологические механизмы, которые как бы «скрыты» от глаз исследователя в норме, но «обнажены» у больных.