


Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
ПАРАМЕТРИЗАЦИЯ СКОРОСТЕЙ ТРАНСФОРМАЦИИ РАСТИТЕЛЬНОГО ОПАДА В ЗАВИСИМОСТИ ОТ ЕГО ХИМИЧЕСКОГО СОСТАВА
Быховец С.С.1, Ларионова А.А.1, Лебедев В.Г.2, Шестибратов К.А.2
1Институт физико-химических и биологических проблем почвоведения РАН, Пущино, Россия.
s_bykhovets@rambler.ru larionova_al@rambler.ru
2Филиал Института биоорганической химии имени академиков М.М. Шемякина и Ю.А. Овчинникова РАН, Пущино, Россия.
vglebedev@mail.ru shestibratov.k@yandex.ru
Аннотация: Рассматриваются возможности уточнения параметризации скоростей трансформации растительного опада в модели ROMUL в зависимости от его химического состава, в частности, содержания азота. Показано, что при повышенных концентрациях азота зависимость скорости минерализации от концентрации ослабевает, а оценка скорости гумификации для древесных остатков требует дальнейшего уточнения, в частности, с учетом содержания лигнина.
Модель динамики органического вещества лесных почв ROMUL широко используется для решения задач лесной экологии и почвоведения. Оценка ее параметров, в частности, скоростей минерализации и гумификации растительных остатков была выполнена на основе как литературных данных, так и лабораторных экспериментов и неоднократно уточнялась (Чертов, 1985; Chertov et al., 2001; Моделирование…, 2007; Комаров и др., 2012). Тем не менее, в рамках работы по оценке возможного влияния предполагаемых плантаций трансгенных форм деревьев на биологический круговорот (Комаров и др., 2013), были начаты новые исследования с целью уточнения указанных параметров в более широком диапазоне концентраций азота и лигнина в опадах.
В работе использованы результаты лабораторных экспериментов по разложению растительных тканей (листья, стволы, корни) молодых растений осины и березы (в т.ч. трансгенных), выращенных в вегетационном опыте. Для оценки параметров модели экспериментальные данные аппроксимировались теоретической кривой, основанной на аналитическом решении соответствующих уравнений модели ROMUL для отдельной порции опада L0 при постоянных коэффициентах:
Y (t) = L(t) + F(t) = L [(1− |
k3 |
|
|
)e−(k1 +k3 )t + |
|
k3 |
|
|
e−k2 |
t ] |
, |
|
0 |
k |
+k |
−k |
2 |
|
k |
+k |
−k |
2 |
|
|
|
|
1 |
3 |
|
|
1 |
3 |
|
|
|
|
||
где Y – “неразложившийся остаток” (оцениваемый по кумулятивной эмиссии углерода), в терминах модели представляющий собой сумму неразложившегося (L) и частично гумифицированного (F) опада, k1, – скорость минерализации пула L, k2 – скорость минерализации пула F, k3 – скорость первичной гумификации (образования F из L), t - время.
Полученные зависимости коэффициентов модели при «стандартных» оптимальных условиях (температура 22°C, влажность субстрата 80% ППВ) от содержания азота (рис. 1) несколько отличаются от полученных ранее, и принятых в современной версии модели ROMUL (Моделирование..., 2007; Комаров и др., 2012). Данные отличия объясняются спецификой анализируемого растительного материала. В частности, расширением диапазона концентраций азота в исследуемом органическом веществе до 3-4% (в ранее исследованных опадах лесных растений обычно не превышали 2%) и большим количеством проб древесины
сразличным содержанием лигнина, ранее авторами модели исследовавшихся мало.
Вчастности, коэффициент k1 в диапазоне концентраций азота 2-4% растет медленнее, чем следует из экстраполяции зависимости, принятой в базовой версии модели. Это достаточно логично, т.к. при высоких концентрациях азот перестает быть основным лимитирующим фактором для сообщества организмов-деструкторов. В то же время, в
27

Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
диапазоне концентраций 1-2% полученные значения коэффициента в среднем близки к «старой» параметризации, а их относительно большой разброс можно объяснить различными свойствами древесины, в частности, различным содержанием лигнина. Таким образом, в качестве первого приближения для оценки значений коэффициента следует использовать «старую» зависимость при концентрациях азота до 1.5%, и новую – при больших концентрациях.
|
|
|
k 1 |
|
|
|
0.0018 |
|
k 2 |
|
|
|
|
0.16 |
|
k 3 |
|
|
|
0.025 |
|
|
|
|
|
y = 0.0003x + 0.0002 |
|
|
|
|
y = 0.0078x + 0.0089 (ROMUL) |
|
|||||
|
|
y = 0.0054x + 0.0005 (ROMUL) |
|
|
0.0016 |
|
|
|
0.14 |
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
R2 |
= 0.3497 |
|
|
|
|
|
y = -0.0193x + 0.0838 |
|
|||||
|
0.02 |
|
|
|
|
|
0.0014 |
|
|
|
|
0.12 |
|
|
||||
-1 |
0.015 |
|
|
|
|
-1 |
0.0012 |
|
|
|
|
|
-1 |
0.1 |
|
|
R2 = 0.3907 |
|
сут. |
|
|
|
|
сут. |
0.001 |
|
|
|
|
|
сут. |
0.08 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
0.0008 |
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
k1 , |
0.01 |
|
|
|
|
k2 , |
|
|
|
|
|
k3 , |
0.06 |
|
|
|
|
|
|
|
|
|
0.0006 |
|
|
|
|
|
|
|
|
|
|||||
|
0.005 |
|
y = 0.0013x + 0.0068 |
|
|
0.0004 |
|
|
|
|
|
|
0.04 |
|
|
|
|
|
|
|
|
|
|
|
|
y = 0.0006 (ROMUL) |
|
|
0.02 |
|
|
|
|
||||
|
|
|
|
R2 = 0.1598 |
|
|
0.0002 |
|
|
|
|
|
|
|
|
|
||
|
0 |
|
|
|
|
|
0 |
|
|
|
|
|
|
0 |
|
|
|
|
|
0 |
1 |
2 |
3 |
4 |
|
|
0 |
1 |
2 |
3 |
4 |
|
0 |
1 |
2 |
3 |
4 |
|
|
|
N, % |
|
|
|
|
|
|
N, % |
|
|
|
|
N, % |
|
|
|
|
|
|
Рисунок 1 - Зависимость скоростей преобразования органического вещества |
|
||||||||||||||
|
|
(коэффициентов k1 - k3 |
модели ROMUL) от исходного содержания азота |
|
||||||||||||||
Коэффициент k2 в современной версии модели принят постоянным. Полученные в настоящем исследовании значения коэффициента обнаруживают слабую положительную зависимость от концентрации азота на фоне довольно большого разброса значений. Это свидетельствует о том что, с одной стороны, при достаточно широком диапазоне свойств исходного органического вещества, на данной стадии его трансформации до некоторой степени сохраняется зависимость от этих свойств, с другой – концентрация азота является не единственным и, возможно, не главным фактором, определяющим данный процесс.
Коэффициент k3 (скорость первичной гумификации) на первый взгляд, обнаруживает зависимость противоположную ранее принятой. Однако можно заметить, что точки в правой части графика, соответствующие листьям, практически идеально ложатся на «старую» прямую, полученную из анализа преимущественно листовых опадов. А наиболее отличающиеся от нее точки в левой части соответствуют одревесневшим побегам (ранее исследованным мало), к тому же, в силу специфики эксперимента, существенно различающимся по биохимическому составу. Таким образом, ранее принятая зависимость k3 от концентрации азота, хорошо работающая для листовых опадов, требует уточнения для древесных. Улучшить данную параметризацию может, в частности, учет концентрации лигнина.
Литература
Комаров А.С., Быховец С.С., Ларионова А.А., Шанин В.Н., Лебедев В.Г., Шестибратов К.А. Модельная оценка биологического круговорота в лесных плантациях с генно-модифицированными деревьями осины // Материалы 3й конференции «Математическое моделирование в экологии» ЭкоМатМод-2013. - Пущино, 2013. - С. 137-138.
Комаров А.С., Хораськина Ю.С., Быховец С.С., Безрукова М.Г., Чертов О.Г. Моделирование динамики органического вещества и элементов почвенного питания в минеральной почве и лесной подстилке // Математическая биология и биоинформатика. - 2012.- Т. 7.- № 1.- С. 162–176.
Моделирование динамики органического вещества в лесных экосистемах / Отв. ред. В.Н. Кудеяров. - М.: Наука,
2007. – 380 с.
Чертов О.Г. Имитационная модель минерализации и гумификации лесного опада и подстилки // Журн. общ. биологии. - 1985. - Т. 46. - № 6. - С. 794-804.
Chertov O.G., Komarov A.S., Nadporozhskaya M.A., Bykhovets S.S., Zudin S.L. ROMUL – a model of forest soil organic matter dynamics as a substantial tool for forest ecosystem modeling // Ecological Modelling. - 2001. - Vol. 138. - P. 289-308.
28

Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
УРАВНЕНИЯ, ОПИСЫВАЮЩИЕ ДИНАМИКУ КОНЦЕНТРАЦИЙ СУБСТРАТА, ПРОМЕЖУТОЧНЫХ И КОНЕЧНЫХ ПРОДУКТОВ, СОДЕРЖАЩИХ 13C, ДЛЯ ИДЕНТИФИКАЦИИ МЕТАБОЛИЧЕСКИХ ПУТЕЙ МЕТАНИЗАЦИИ ЦЕЛЛЮЛОЗЫ
Вавилин В.А., Рытов С.В.
Институт водных проблем РАН, Москва, РФ vavilin@aqua.laser.ru
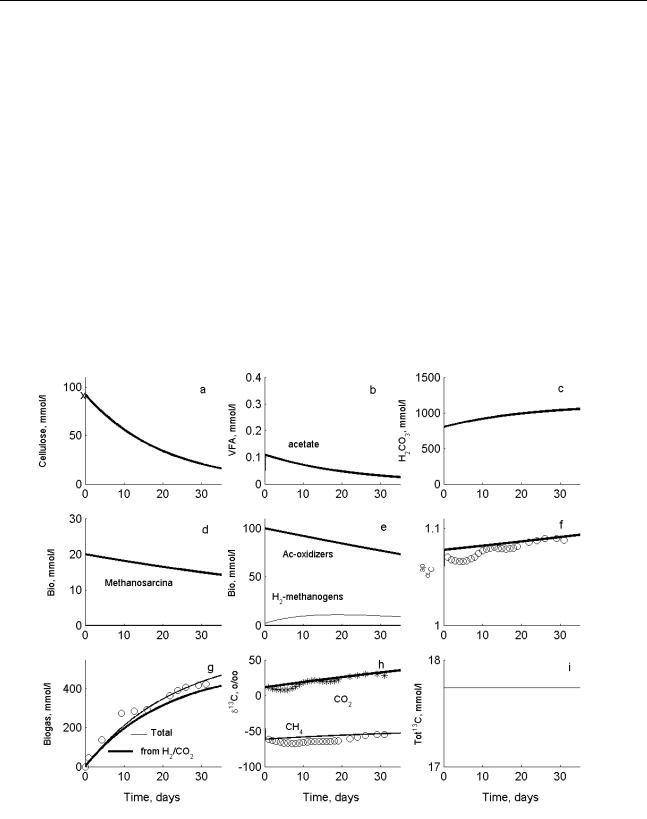
Аннотация: Традиционные уравнения, игнорирующие изотопный состав, дополняются дифференциальными уравнениями, описывающими динамику концентраций тяжелых компонентов, содержащих изотоп 13С (субстрат, промежуточные и конечные продукты, микроорганизмы). При разложении целлюлозы идет синтрофное окисление ацетата, сопровождаемое значительным повышением содержания 13С в СО2, в свою очередь, приводящим к возрастанию эмпирического коэффициента фракционирования изотопов углерода между CO2 и
CH4
Целюлоза и гемицеллюлоза являются основными разлагаемыми компонентами в естественных и некоторых технологических системах. В ходе метаногенеза целлюлозного материала участвуют многочисленные микроорганизмы, осуществляющие деполимеризацию (гидролиз), ферментативный кислотогенез, ацетогенез и метаногенез. Основными субстратами для ацетикластических и гидрогенотрофных метаногенных микроорганизмов являются ацетат и водород/диоксид углерода:
CH3COOH + H 2O → CH 4 + H 2CO3 |
(1) |
4H 2 + H 2CO3 → CH 4 + 3H 2O |
(2) |
Традиционно считается, что вклад ацетикластического метаногенеза составляет около 70% общей продукции метана. Однако, относительный вклад различных ацетикластических и гидрогенотрофных метаногенных микроорганизмов в образование метана может определяться факторами окружающей среды, типом анаэробного реактора и режимом проведения процесса, а также составом микроорганизмов в инокуляте. В присутствии ингибиторов, таких как аммоний и летучие жирные кислоты, доминирующим механизмом метанообразования является процесс, при котором ацетат предварительно окисляется до H2 и H2CO3:
CH3COOH + 4H2O → 4H2 + 2H2CO3 |
(3) |
Эта реакция происходит синтрофно с гидрогенотрофными метаногенами, превращающими
H2 и H2CO3 в CH4.
Поскольку величина отношения стабильных изотопов 13С/12С составляет порядка 0,01, она традиционно определяется при помощи величин “приращения ”, которые выражаются в промилях o / oo разницы от стандарта PDB:
δ!3C(o / oo) = [ |
(13C/12C)îáð |
(4) |
|
|
−1]×1000 |
||
(13C/12C)PDB |
|||
где индекс “обр” - образец, PDB - стандарт с высоким содержанием 13С в карбонате |
|||
кальция особого моллюска. Для уточнения доминирующего пути превращения |
|
||
целлюлозы в метан часто применяется эмпирический фактор фракционирования |
|
||
изотопов между CO2 и CH4: |
|
||
αcapp = δ13CCO2 |
+1000 |
|
(5) |
δ13CCH4 |
+1000 |
|
|
29
Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
Ранее (Вавилин, Рытов, 2015), для описания динамики изотопа 15 N в процессе превращения нитрата (субстрат) в закись азота (продукт) применялись следующие уравнения:
d15NO3- |
|
|
15 |
NO3- |
1 |
|
− |
|
|||
dt |
|
= − |
|
|
|
|
|
ρ(NO3 ,Org,B) |
|
||
|
NO3- |
|
α N |
|
|||||||
|
|
|
|
|
|
|
|||||
d15N14 NO |
= − |
1 |
(1−Y − ) |
d15NO3- |
(6) |
||||||
dt |
|
|
2 |
dt |
|||||||
|
|
|
|
|
|
NO3 |
|
||||
|
|
|
|
|
|
|
|
|
|
||
d 15NB |
= |
dB 15 NO3- |
1 |
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
dt |
dt NO3- |
|
α N |
|
|
|
|||||
|
|
|
|
|
|||||||
где ρ( NO3− , Org, B) - скорость процесса как функция от концентрации нитрата ( NO3− ), органического материала (Org) и денитрифицирующей биомассы (B); 15 NO3− , 15 N14 NO , 15 B - концентрации тяжелых компонентов, соответственно; YNO3− - экономический коэффициент,
αN - фактор фракционирования изотопов. Эти уравнения послужили основой для описания концентраций тяжелых составляющих в ходе разложения целлюлозы.
В итоге было показано, что динамика αCap определяет доминирующий путь разложения целлюлозы, а именно, синтрофное окисление ацетата.
Литература
Вавилин В.А., Рытов С.В. Нелинейная динамическая модель фракционирования стабильных изотопов азота в процессе денитрификации с образованием закиси азота // Вод. Рес. - 2015. - № 2. - С.194-198.
30