


Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
ИЗУЧЕНИЕ МЕХАНИЗМОВ КОНКУРЕНЦИИ В СМЕШАННЫХ ДРЕВОСТОЯХ С ПОМОЩЬЮ ИМИТАЦИОННОЙ МОДЕЛИ С АДАПТИВНЫМ АЛГОРИТМОМ
Шанин В.Н. 1, Шашков М.П. 1, Иванова Н.В. 2, Рочева Л.К. 3, Москаленко С.В. 1, Бурнашева Э.Р. 4, Комаров А.С. 1, Mäkipää R. 5
1Институт физико-химических и биологических проблем почвоведения РАН, г. Пущино, Российская Федерация
shaninvn@gmail.com
2Институт математических проблем биологии РАН, г. Пущино, Российская Федерация 3Администрация ГО Пущино, г. Пущино, Российская Федерация 4Башкирский Государственный Университет, Уфа, Российская Федерация
5Natural Resources Institute Finland, Vantaa, Finland
Аннотация: Предложена новая модель корневой конкуренции между деревьями в смешанных древостоях, учитывающая вертикальную и горизонтальную неравномерность в распределении корней древесных растений разных видов и варьирование условий местообитания.
1.Описание модели
Впредложенной модели имитируемое пространство представлено квадратной решеткой, разделенной на ячейки одинакового размера. Площадь зоны питания каждого дерева зависит от диаметра ствола, вида дерева и условий местообитания. Очередность включения новых ячеек в зону питания зависит от расстояния между текущей ячейкой и базовой (т.е., той, в которой располагается само дерево), массы корней других деревьев в текущей ячейке и количества доступных элементов питания (прежде всего, азота) в данной ячейке. Согласно формулировке модели, элементы минерального питания в каждой ячейке распределяются между деревьями пропорционально биомассе их тонких корней в данной ячейке, с дополнительными поправками на расстояния до ствола, вид и возраст дерева (Лебедев, 2012; Лебедев, Лебедев, 2012). Модель описывает вертикальное распределение биомассы корней по профилю почвы посредством степенной функции с видоспецифичными коэффициентами. Скорость оборота тонких корней неодинакова в пределах зоны питания и определяется отношением потребленного на текущем шаге азота к массе тонких корней в данной ячейке.
2.Результаты и обсуждение
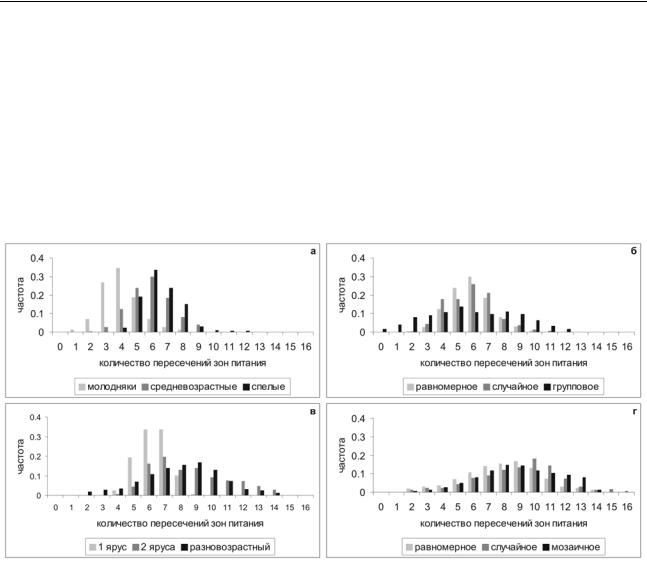
Интенсивность корневой конкуренции возрастает от молодых древостоев к спелым. Также в этом ряду возрастает однородность в распределении подземной биомассы. При этом для древостоев всех возрастных групп распределение плотности пересечений зон питания имеет колоколообразный вид (рисунок 1а).
Различия в характере начального расположения деревьев также оказывали влияние на характер конкуренции. При регулярном расположении (по квадратной решетке) и при случайном расположении деревьев обычно наблюдалось колоколообразное распределение плотности пересечений зон питания (с незначительными отклонениями во втором случае), в то время как при групповом расположении деревьев (в нескольких плотных кластерах) распределение пересечений зон питания имело бимодальный характер. Правая часть данного распределения соответствует конкуренции внутри кластеров, тогда как левая часть – «окнам» между кластерами, где интенсивность конкуренции невысока (рисунок 1б).
В одновозрастных древостоях, по сравнению с древостоями с более сложной структурой, количество пересечений зон питания было ниже. Кроме того, для древостоя с двумя древесными ярусами наблюдалось бимодальное распределение (рисунок 1в).
195
Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
Различный характер пространственного распределения азота в доступной для растений форме также оказал влияние на характер распределения подземной биомассы в древостое. Случайное и мозаичное распределение привело к усилению корневой конкуренции (гистограмма скошена вправо, среднее количество пересечений выше) по сравнению с однородным распределением. Правая часть гистограмм, скорее всего, соответствует распределению корней в богатых азотом участках имитационной решетки, где корневая конкуренция выше (рисунок 1г).
Результаты моделирования показали, что увеличение сложности структуры древостоев (в т.ч. пространственное расположением деревьев, видовой состав, возрастная структура, вертикальная структура) выражается в более сложном пространственном характере конкурентных взаимоотношений.
Рисунок 1 – Распределение интенсивности корневой конкуренции (количество пересечений зон питания на ячейку имитационной решетки) для (а) древостоев разного возраста (на примере одновидовых древостоев со случайным расположением деревьев); (б)
при различном характере расположения деревьев (на примере средневозрастных одновидовых древостоев); (в) при различной вертикальной структуре древостоев (средневозрастный березняк, березняк с подростом ели, разновозрастный ельник); (г) при разном характере пространственного распределения доступных соединений азота в почве – равномерно по всем ячейкам имитационной решетки, случайно с варьированием в 20% от среднего, крупными «пятнами» с варьированием в 20% от среднего (на примере разновозрастного ельника). Для всех случаев, кроме (г) полагалось, что соединения азота в почве равномерно распределены по всем ячейкам имитационной решетки.
Благодарности
Работа выполнена при финансовой поддержке РФФИ (грант № 15-04-05400) и Академии наук Финляндии (гранты №№ 140766 и 278151).
Литература
Лебедев Е.В. Продуктивность берёзы белой на уровне организма в онтогенезе в европейской части России // Известия Оренбургского государственного аграрного университета. – 2012. - № 4. - С. 18-22.
Лебедев В.М., Лебедев Е.В. Взаимосвязь биологической продуктивности и поглотительной деятельности корней хвойных пород в онтогенезе в зоне южной тайги России // Агрохимия. – 2012. - № 8. - С. 9-17.
196

Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
ИЗМЕНЕНИЯ ОРГАНИЧЕСКОГО ВЕЩЕСТВА В ЛЕСНЫХ ПОЧВАХ ПОД ПЛАНТАЦИЯМИ С БЫСТОРОРАСТУЩИМИ ФОРМАМИ БЕРЕЗЫ
Шанин В.Н. 1, Комаров А.С. 1, Шестибратов К.А. 2
1Институт физико-химических и биологических проблем почвоведения РАН, г. Пущино, РФ shaninvn@mail.com; as_komarov@rambler.ru
2Филиал Института биоорганической химии имени академиков М.М. Шемякина и Ю.А. Овчинникова РАН, г. Пущино, РФ
schestibratov.k@yandex.ru
Аннотация: сформулирована задача анализа ускоренного круговорота органического вещества и азота в планируемых плантациях быстрорастущих биотехнологических форм березы; приводится пример анализа этой ситуации с помощью системы моделей роста леса и биологического круговорота EFIMOD; показана необходимость применения органических удобрений для сохранения баланса углерода и азота в таких плантациях.
При создании промышленных лесных плантаций, заложенных на основе быстрорастущих древесных видов с улучшенными ростовыми характеристиками и биохимическим составом, неизбежно возникают вопросы о влиянии таких растений на окружающую среду. С целью анализа возможных последствий выращивания модифицированных растений березы (Betula pendula Roth), с учетом данных, полученных в ФИБХ РАН (Shestibratov et.al., 2011),) для оценки потенциала накопления биомассы и динамики запасов азота в почве в плантациях биотехнологических форм березы по сравнению с нативной формой была использована система моделей EFIMOD (Komarov et al., 2003).
Компьютерный эксперимент предусматривал имитацию динамики древостоя и почвы на березовых плантациях в течение 5 оборотов рубки; продолжительность оборота рубки составила 25 лет. Во время первой из имитируемых ротаций происходила автоподстройка модели, поэтому данная ротация не учитывалась при последующем анализе.
Были использованы 3 имитационных сценария: (1) выращивание плантаций нативных форм березы; (2) выращивание плантаций быстрорастущих форм березы без удобрений и (3) выращивание плантаций быстрорастущих форм березы с внесением удобрений. При параметризации модели было принято допущение, что быстрорастущие формы березы имеют на 25% большую максимальную биологическую продуктивность (Shestibratov et.al., 2011).
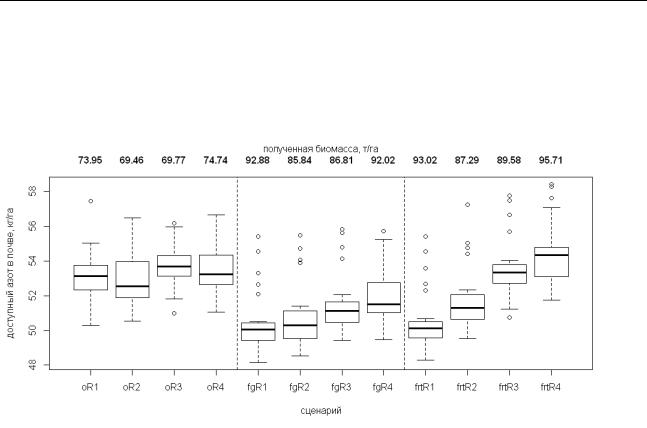
В пределах каждого оборота рубки имитировались следующие лесохозяйственные мероприятия: (1) внесение органических удобрений на поверхность почвы перед посадкой (только для сценария (3)); (2) посадка саженцев березы; (3) сплошная рубка при возрасте древостоя 30 лет. Предполагалось, что массовая доля азота в удобрении составляет 0.35%, а норма внесения – 3 т га-1 (Желдак, Атрохин, 2002). Посадка березы имитировалась по регулярной решетке с расстоянием между рядами в 5 м. Высота саженцев составляла 1.5±0.5 м, диаметр на уровне груди 0.75±0.025 см; данные параметры были определены на основе региональных таблиц хода роста (Швиденко и др., 2008). Моделирование рубки предполагало, что с лесосеки изымается 100% массы стволов, 50% массы ветвей и 10% массы листвы. В качестве контролируемых переменных были выбраны: (1) общее количество доступного азота в почве и (2) растительная масса, полученная на каждом обороте рубки. Результаты моделирования (Рисунок 1) показали, что плантации биотехнологических форм демонстрируют более интенсивный рост и накопление фитомассы, что, в свою очередь, приводит к более интенсивному потреблению деревьями соединений азота из почвы. Внесение удобрений позволяет в значительной степени
197
Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
компенсировать усиленное потребление элементов минерального питания. Кроме того, к концу четвертого оборота рубки, наблюдается рост количества доступных соединений азота в почве. Данный факт может объясняться тем, что быстрорастущие формы березы обеспечивают более высокое, по сравнению с нативными формами, поступление растительного опада, который вовлекается в процессы минерализации в почве. Одним из конечных продуктов минерализации являются неорганические соединения азота.
Рисунок 1. Результаты моделирования по выбранным сценариям: полученная в ходе лесозаготовки биомасса деревьев, т/га (верхний ряд чисел) и динамика содержания почвенного азота в форме доступных для растений соединений, кг/га, за один оборот рубки, т.е. за 25 лет (приведены медианное значение, 2-й и 3-й квартили и размах; малыми окружностями показаны «выбросы»). Обозначения сценариев: «o» – плантации нативной формы березы; «fg» – плантации быстрорастущей формы березы, сценарий без внесения удобрений; «frt» – плантации быстрорастущей формы березы, сценарий с внесением удобрений.
Благодарности
Работа выполнена при поддержке Министерства образования и науки РФ рамках проекта № 14.616.21.0013 от 17 сентября 2014 года (уникальный идентификатор RFMEFI61614X0013) по теме: «Разработка технологии оценки и прогнозирования экологических эффектов выращивания лесных плантаций на основе биотехнологических форм деревьев с заданными свойствами (увеличенная скорость роста, усиленная ассимиляцией азота почвы, пониженное содержание лигнина, повышенное содержание целлюлозы)».
Литература
Желдак В.И., Атрохин В.Г. Лесоводство, ч. I. – М.: ВНИИЛМ, 2002. – 336 с.
Швиденко А.З., Щепащенко Д.Г., Нильссон С., Булуй Ю.И. Таблицы и модели роста и продуктивности основных лесообразующих пород Северной Евразии (нормативно-справочные материалы). – М.: Федеральное агентство лесного хозяйства. Международный институт прикладного системного анализа,
2008. – 886 с.
Komarov A.S., Chertov O.G., Zudin S.L., Nadporozhskaya M.A., Mikhailov A.V., Bykhovets S.S., Zudina E.V., Zoubkova E.V. EFIMOD 2 – a model of growth and cycling of elements in boreal forest ecosystems // Ecol. Mod. - 2003. - V. 170. - P. 373–392.
Schestibratov K., Lebedev V., Podrezov A., Salmova M. Transgenic aspen and birch trees for Russian plantation forests // BMC Proceedings. - 2011. - Vol. 5 (Suppl. 7). - P. 124.
198