

Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
ПЕРЕСТРОЙКА СТРУКТУРЫ СООБЩЕСТВА МЕЛКИХ МЛЕКОПИТАЮЩИХ – ПРЕДВЕСТНИК ИЗМЕНЕНИЯ РЕЖИМА ДИНАМИКИ ЧИСЛЕННОСТИ?
Кшнясев И.А., Давыдова Ю.А.
Институт экологии растений и животных УрО РАН, Екатеринбург, Россия kia@ipae.uran.ru
Аннотация: Исследована многолетняя (1982–2014) динамика плотности населения и структуры сообщества мелких млекопитающих (СММ) в южно-таежных лесах. Показаны существенные перестройки СММ в ответ на нарушения (вывал леса, пожар и последующие сукцессии) биоценозов при сохранении трехлетней цикличности и последующее «спонтанное» изменение режима – переход к квазидвухлетней периодичности. Предложен сценарий, объясняющий различный наблюдаемый режим флуктуаций.
Различный режим (амплитуда, период) динамики популяций грызунов в географическом градиенте или его изменение в фиксированной точке приписывается
(Hansson, Henttonen, 1985; Кшнясев, 2001; Hörnfeldt, 2004; Cornulier et al., 2013; Korpela et al., 2014) эффектам внешних факторов – особенностям: климата (и/или его изменению), структуры сообщества хищников (соотношению специалистов/генералистов). Мы высказываем (основанное на наших наблюдениях) предположение, что не менее правдоподобна и противоположная версия – влияние изменений условий среды вследствие внешних воздействий (как аналога климатического градиента) может быть опосредовано (усилено или, наоборот, демпфировано) перестройкой структуры СММ. Изменение «композиции» СММ вследствие особенностей (трофической специализации, различного репродуктивного потенциала, привлекательности для хищника и др.), составляющих его видов способно порождать заметные сдвиги внутри- и межвидовой конкуренции в СММ и взаимодействий со специализированными хищниками – миофагами, и далее отражаться в сохранении или vice versa в изменении режима динамики численности.
В мониторинговых исследованиях (по контрасту с управляемым экспериментом) главный вызов (или «логическая ловушка») для интерпретатора – отделение симптомов от причин. Некоторую надежду на «объективность» теории (на фоне априорных представлений и прошлого опыта) дает рассмотрение результатов BACI – «натурных экспериментов» – реакции (как подвергнутых воздействию – I (impact), так и интактных – C (control) сообществ на нарушения (B – before A – after), и последующую интерпретацию в духе “post hoc, ergo propter hoc”.
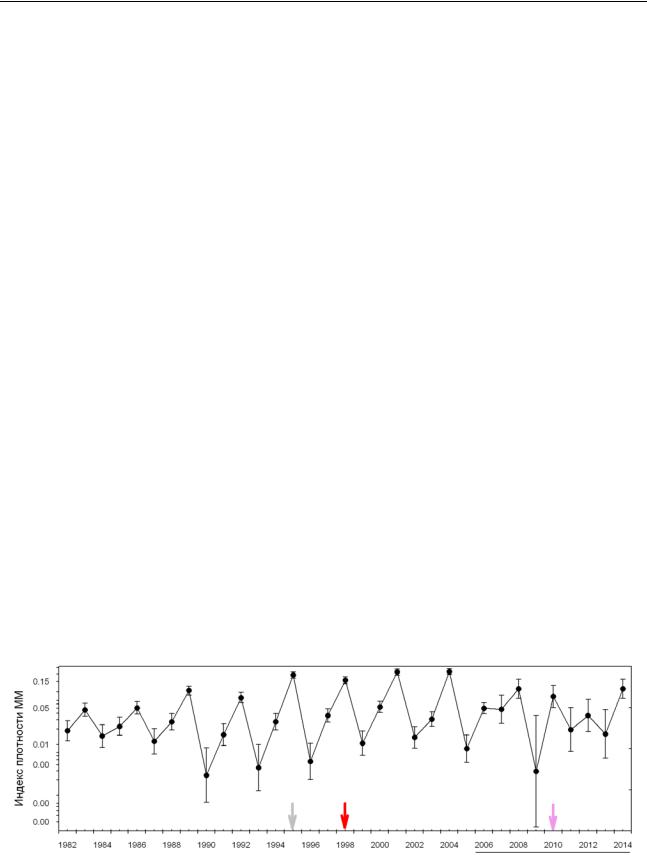
Рисунок 1. Плотность населения СММ весной (p=n/N, 95% ДИ, логит шкала). Стрелки –
нарушения: вывал леса (1995), пожар (1998, 2010); подчеркнутые годы – утрата трехлетней периодичности. Средний Урал, южная тайга, Висимский заповедник; 1982-2014.
95
Материалы Четвертой конференции «Математическое моделирование в экологии» ЭкоМатМод-2015, г. Пущино, Россия
Динамика плотности населения СММ (рисунок 1) до 2006 г. может быть представлена (если опустить непостоянство амплитуды) как простой трехлетний цикл, и изменения ее режима не выявляется. Однако в композиции СММ четко обнаруживается отклик (идеально согласующийся с представлениями об экологических нишах видов) на изменения условий среды (рисунок 2). Перестройке структуры СММ можно приписать и последующее изменение динамики численности населения, вследствие неидентичности репродуктивного потенциала, особенностей внутри- и межвидовой конкуренции (асимметрия и «порог включения»). Например, после 2004 года, мы уже не наблюдали (ранее весьма специфичного «симптома» фазы пика) блокировки созревания сеголеток у доминирующего вида – рыжей полевки.
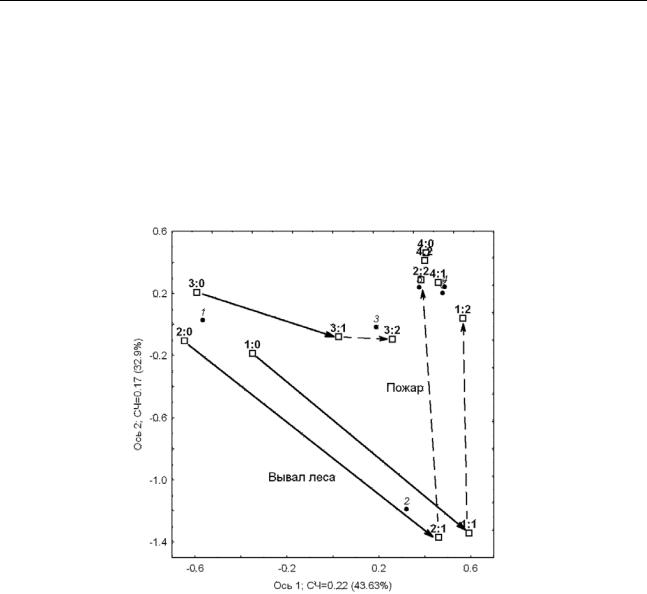
Рисунок 2. Перестройка структуры сообщества ММ. Результаты ординации матрицы
6×12: «виды»(6)×время(3: 19820–19951 –19982 –2006)-участки(4), проекция на плоскость
двух первых канонических осей. Виды×Время: X2(10)=1647.27.
Точки – виды ММ (1-3 – лесные и 4 – серые полевки, 5 – бурозубки, 6 – лесная мышь); квадраты – участки: 1 и 2 – коренной лес в горной части, 3 – вторичный лес на равнине, 4
– зарастающая лесом елань; до нарушения (:0) → после вывала леса (:1) → после пожара (:2). Средний Урал, южная тайга, Висимский заповедник, 1982-2006.
Литература
Кшнясев И.А. Одна популяция – два режима динамики? // Современные проблемы популяционной, исторической и прикладной экологии: Мат. конф. Вып. 2. - Екатеринбург, 2001. - С. 132-137.
Cornulier T., and 20 other. Europe-wide dampening of population cycles in keystone herbivores // Science. - 2013. - № 340. - P. 63-66.
Hansson L., Henttonen H. Gradients in density variations of small rodents: the importance of latitude and snow cover // Oecologia. - 1985. - V. 67. № 3. - P. 394-402.
Hörnfeldt B. Long-term decline in numbers of cyclic voles in boreal Sweden: Analysis and presentation of hypotheses // Oikos. - 2004. - V. 107. № 2. - P. 376-392.
Korpela K., and 9 other. Predator–vole interactions in northern Europe: the role of small mustelids revised // Proc. R. Soc. B. - 2014. http://dx.doi.org/10.1098/rspb.2014.2119
96